Экспозиция "Образ жизни некоторых современных и ископаемых организмов"
Класс Двустворчатые моллюски (Bivalvia)
Практически все двустворчатые моллюски - донные, относительно малоподвижные животные. Двустворки распространены в морских, пресноводных и солоноватоводных бассейнах и встречаются на всех глубинах, всех широтах и всех грунтах. По типу питания практически все эти моллюски относятся к сестонофагам (фильтраторам), к сортирующим детритофагам (инфаунным и эпифаунным), и лишь некоторые немногочисленные виды приспособлены к питанию посредством симбиоза с одноклеточными водорослями зооксантеллами (например, гигантские обитатели рифов - моллюски рода Tridacna, в мантии которых поселяются эти водоросли). В периодически осыхающих бассейнах (например, в зоне литорали) двустворки долгое время могут находиться без воды. Среди двустворчатых моллюсков очень много ископаемых форм.
Большая часть двустворок существует на небольших глубинах в тропических и субтропических зонах, в том числе и на рифах при высоких среднегодовых температурах. Для тропических форм характерна яркая окраска и богатая скульптура раковин. Толщина раковины связана с глубиной существования: толстой раковиной обладают мелководные формы, живущие в условиях сильного волнового воздействия, тонкая раковина - у обитателей глубоководных морских и пресноводных водоемов (в таких условиях извлечение извести из воды затруднено). Для мелководных форм важна хорошая аэрация, глубоководные менее чувствительны к этому фактору. Среди двустворок есть как стеногалинные (нормальноморские), так и эвригалинные (обычно солоноватоводные) формы (например, Dreissena, Mactridae).
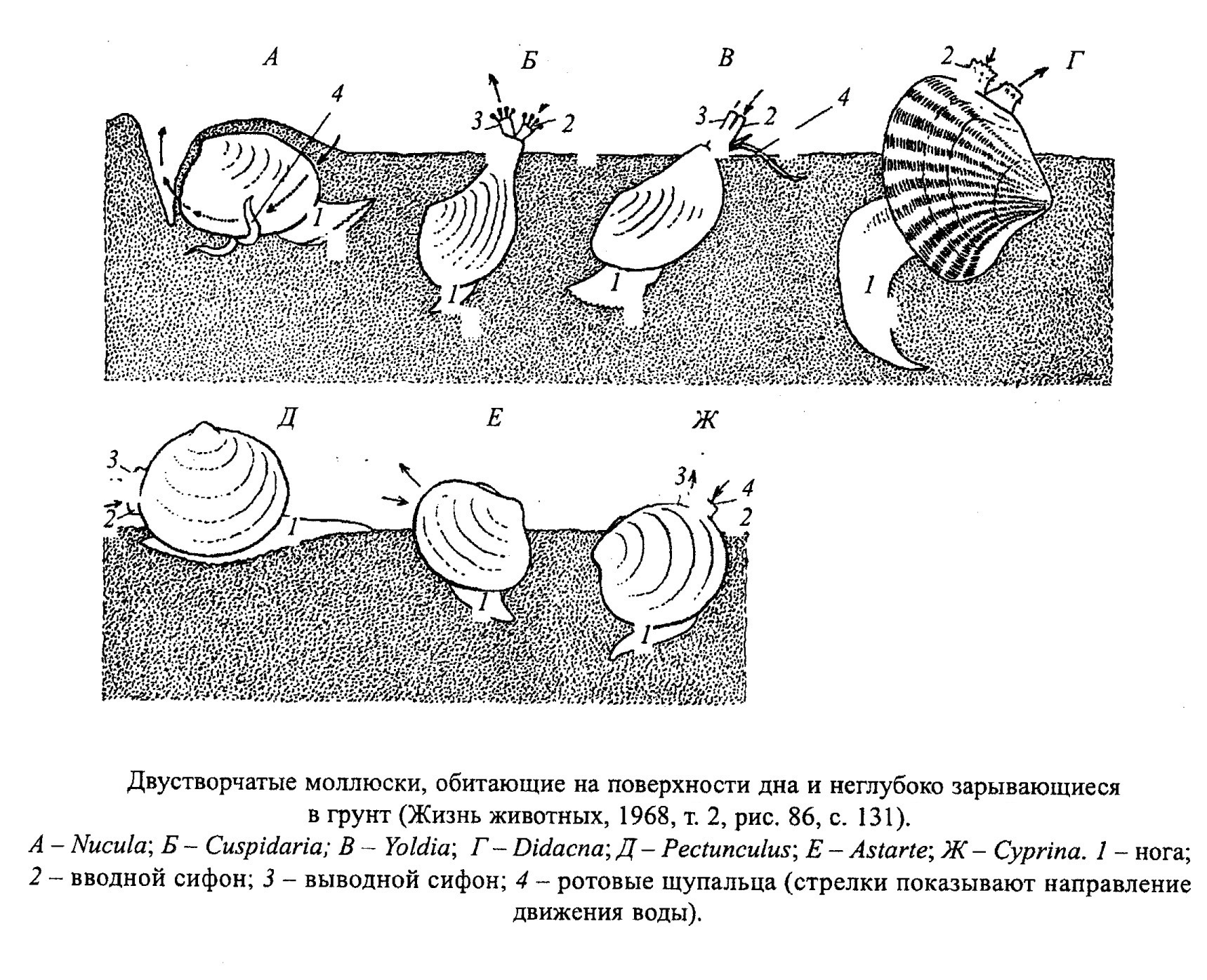
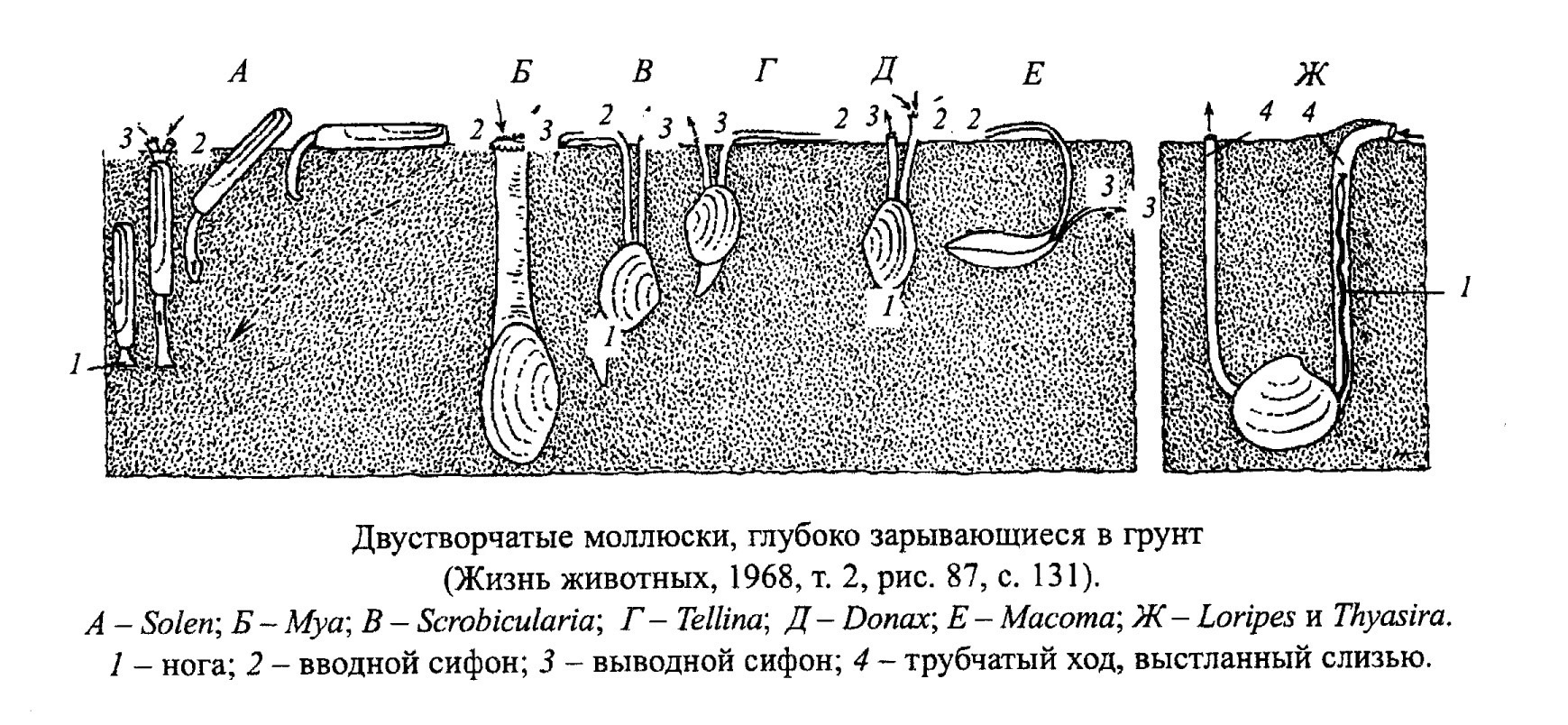
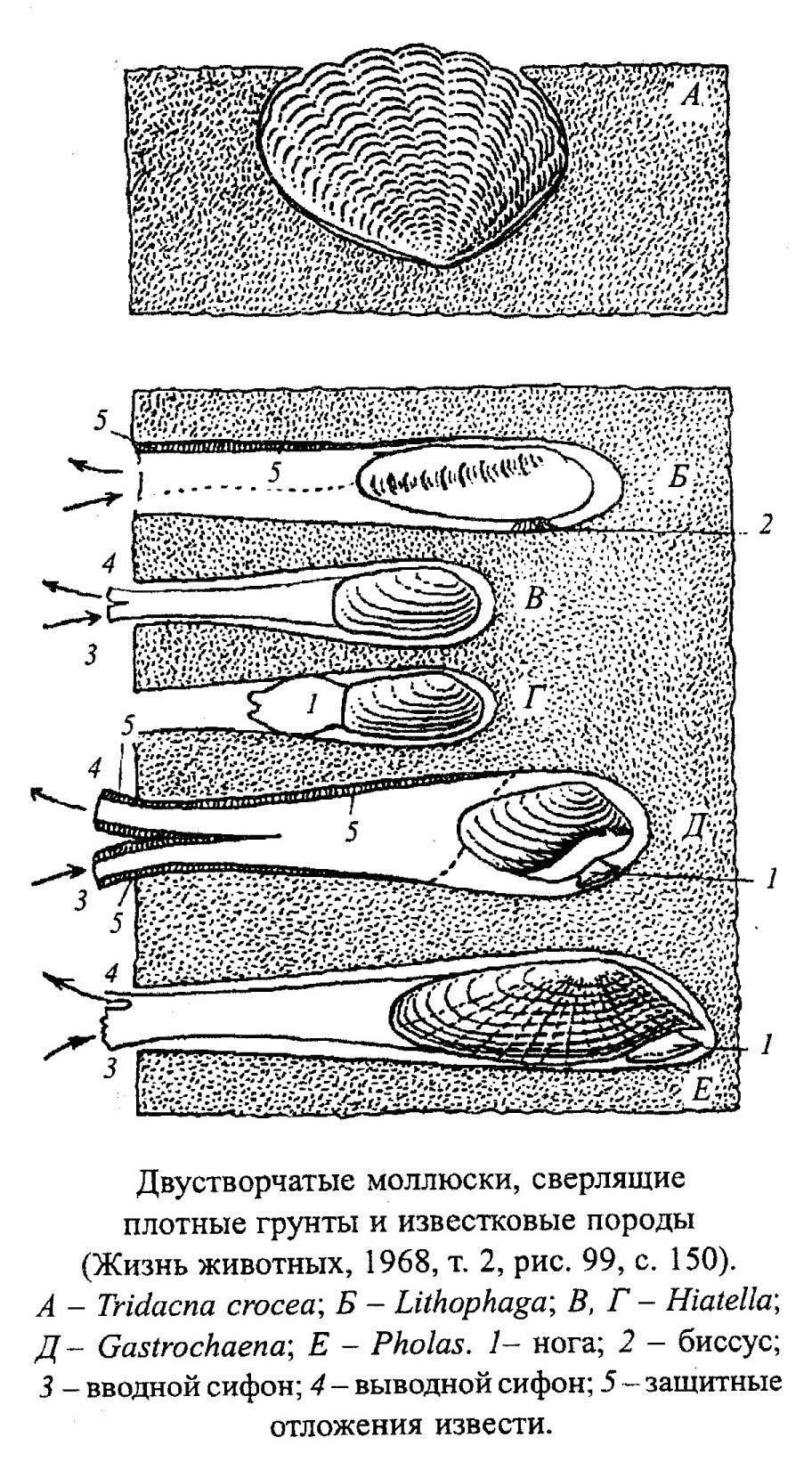
Образ жизни двустворок очень разнообразен: одни прикрепляются биссусом или цементом, другие передвигаются (ползают, прыгают, плавают, зарываются в грунт или сверлят твердый субстрат) (рис. 32, 33, 34, 35, 36, 37).
У разных систематических групп двустворчатых моллюсков, которые, однако, ведут сходный образ жизни и существуют в близких зонах, выработались близкие морфологические особенности. Так, у ползающих обычно равные створки; у лежащих на одной створке нижняя всегда больше; биссусные формы обычно резко несимметричные и неравностворчатые; сверлильщики имеют заостренный передний конец раковины; у глубоко зарывающихся форм обычно тонкая гладкая раковина.
Образ жизни двустворок сильно отличается в зависимости от отношения их к субстрату. Двустворки:
- ползают по дну (беззубки Anodonta, обр. 385-68, Astartidae);
- прикрепляются с помощью биссуса (при этом, например, мидии Mytilidae неподвижно прикрепляются к выступам твердого субстрата в течение всей жизни, обр. 385-51; Dreissena может менять место обитания, обр. 385-52; Arca прикрепляется на ранней стадии развития, а потом свободно перемещается, прикрепляясь лишь периодически; Pinna погружается передним концом в грунт и закрепляется в нем мощным биссусом; Lima, Limatula могут плавать, ползать, зарываться и строить "гнездо", обволакивая частицы грунта биссусом; морской гребешок Pecten на ранних стадиях развития прикрепляется биссусом, а потом плавает брюшным краем вперед, хлопая створками, или свободно лежит на дне);
- свободно лежат на дне (Tridacna, обр. 385-56, Inoceramus в течение всей жизни, обр. 385-54, а некоторые устрицы, например, Exogyra, лишь на взрослой стадии, обр. 385-55);
- прикрепляются с помощью цемента (многие устрицы Ostrea, обр. 385-57, Anomia и др.; толстозубые моллюски рудисты, обр. 385-60);
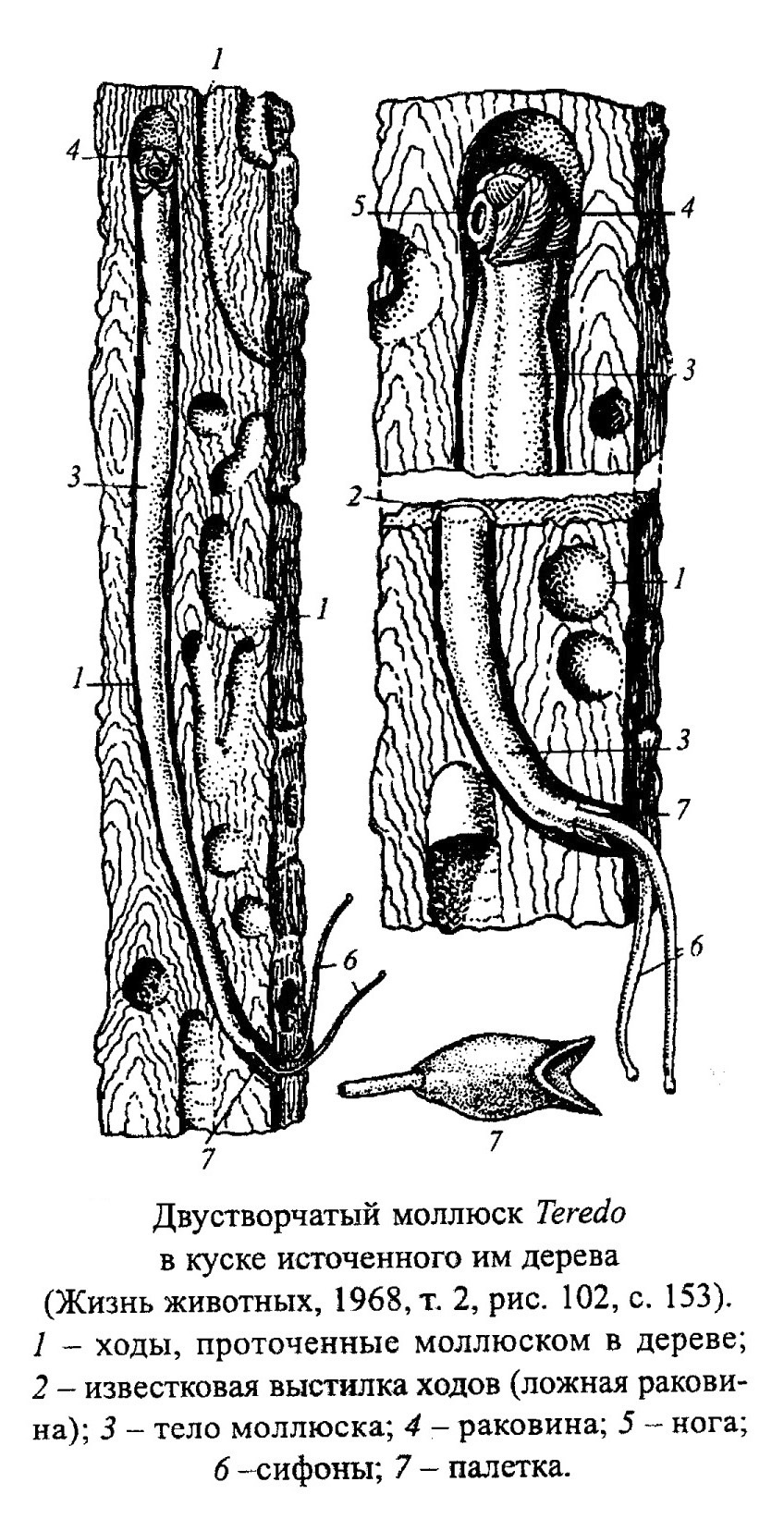
- сверлят твердый субстрат - скальный известняк, раковины других моллюсков, колонии кораллов, древесину (механическим путем сверлят Pholadidae, Theredidae, химическим - Petricola, Lithophaga, обр. 385-62, 385-64).
Двустворчатые моллюски, неподвижно лежащие на дне, не выносят заноса рыхлым осадком, для подвижных форм этот фактор среды не имеет большого значения. Цементаторы, сверлильщики и формы, прикрепленные биссусом к неподвижным предметам, обычно заселяют зону активной гидродинамики.
Двустворки имеют огромное породообразующее значение. В палеозое, мезозое и особенно в кайнозое широко распространены толщи ракушняков, образованные раковинами этих моллюсков.
Класс Головоногие моллюски (Cephalopoda)
Головоногие моллюски - обитатели теплых и умеренных морских бассейнов с нормальной соленостью, очень требовательные к среде обитания организмы.
Современные головоногие, которые обладают раковиной, выполняющей функцию жидкостно-газового гидростатического аппарата (наутилусы, каракатицы), обитают на сравнительно небольших глубинах. Каракатицы не опускаются ниже 250-300 м, наутилусы - ниже 600 м (этот предел ограничивается механической прочностью их раковин). Современные кальмары и осьминоги, не обладающие жидкостно-газовой гидростатикой, распространяются до глубин 5000 м.
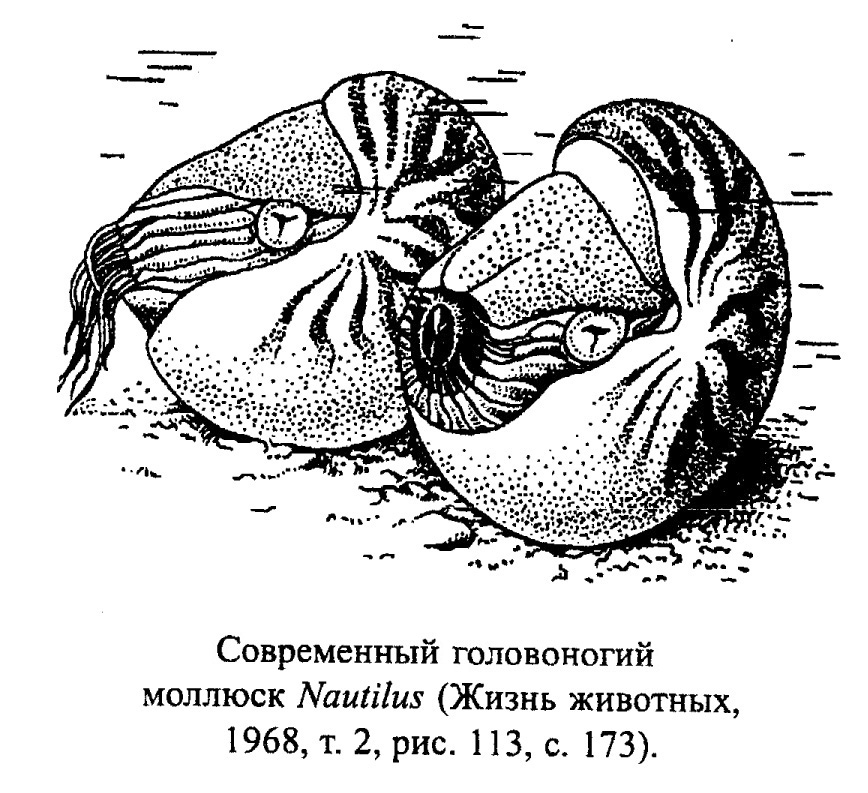
Современные представители подкласса Nautiloidea распространены в тропиках. Это хищники и падальщики, хорошие пловцы, которые, однако, ведут в основном придонный образ жизни, так как питаются различным бентосом (чаще ракообразными), захватывая добычу с помощью многочисленных щупальцев. После гибели некоторых из этих моллюсков их раковины, заполненные газом, всплывают и относятся течениями на значительные расстояния. Так, раковины моллюска Nautilus (рис. 38) находят на Мадагаскаре и в Японии, тогда как его прижизненный ареал лежит между Австралией, Филиппинами и островами Фиджи.
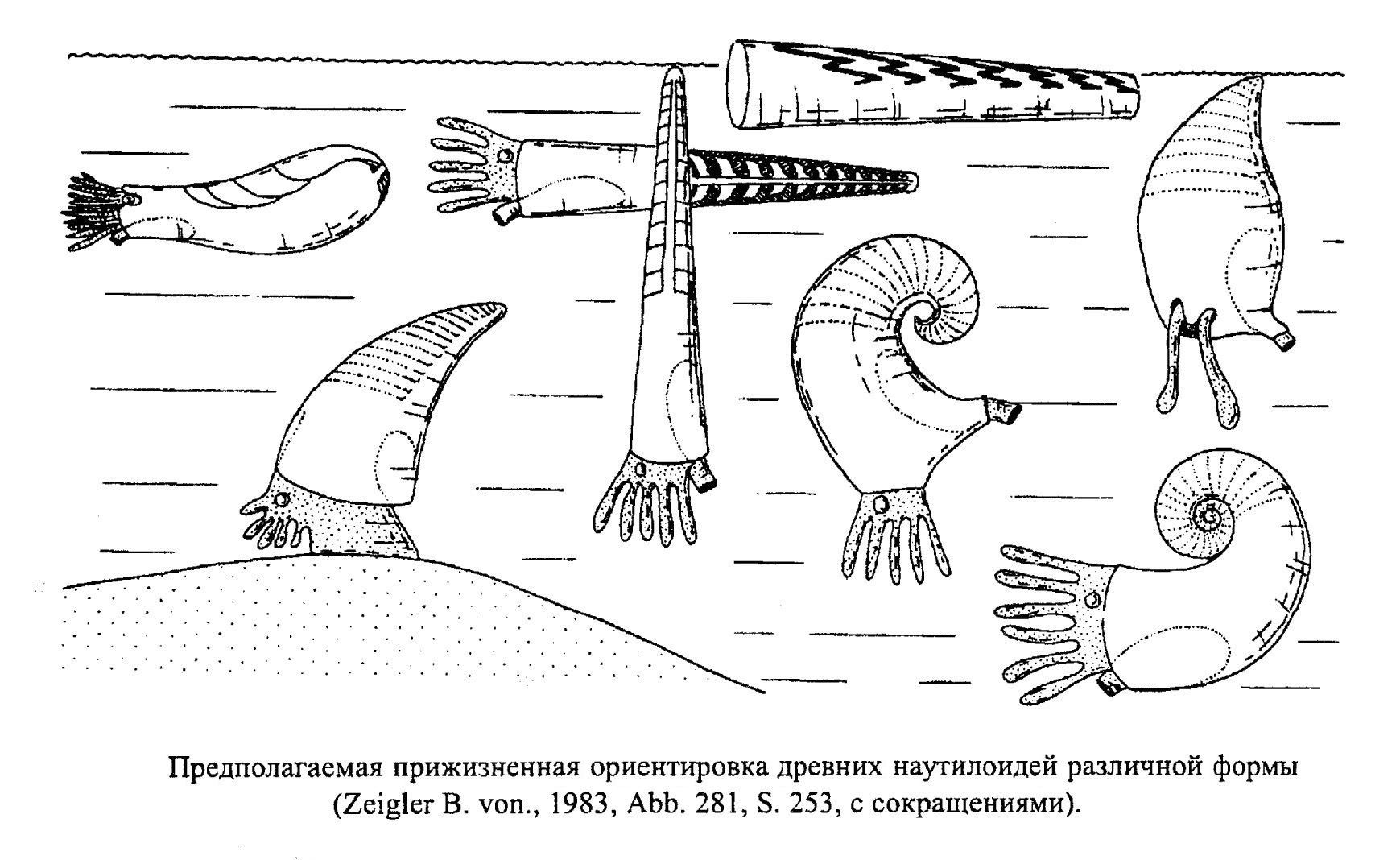
Палеозойские наутилоидеи с прямой раковиной (Orthoceras, Endoceras, обр. 385-96) принадлежали к группе медленно ползающих донных животных. Обладатели спиральной раковины могли не только ползать, но и медленно плавать в горизонтальном и вертикальном направлении. Обычно остатки палеозойских наутилоидей встречаются в отложениях мелководных бассейнов, реже - в пелагических отложениях (рис. 39).
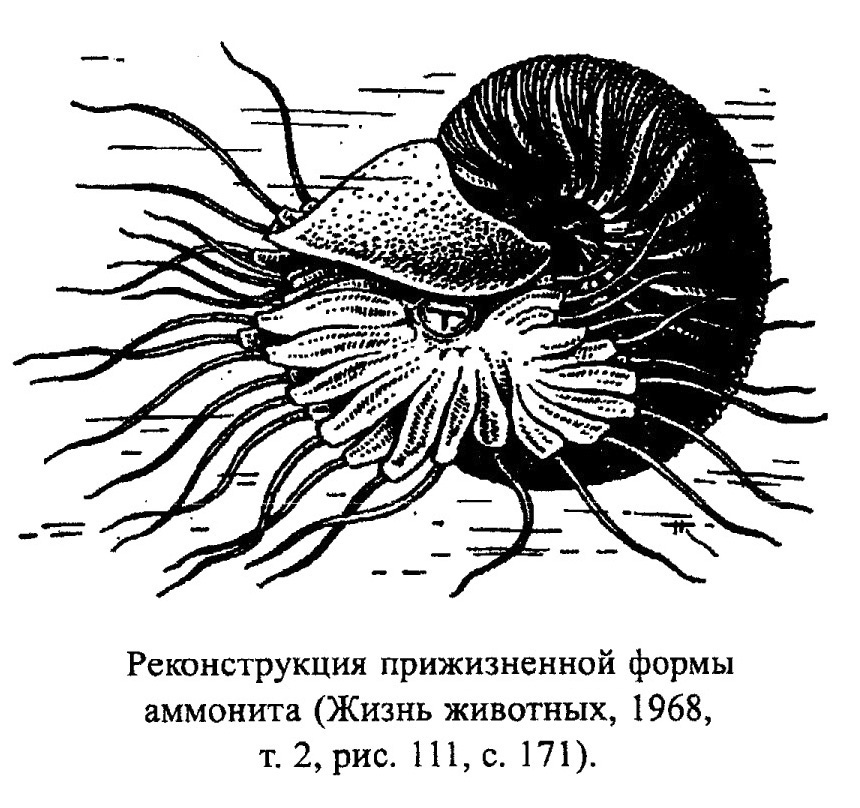
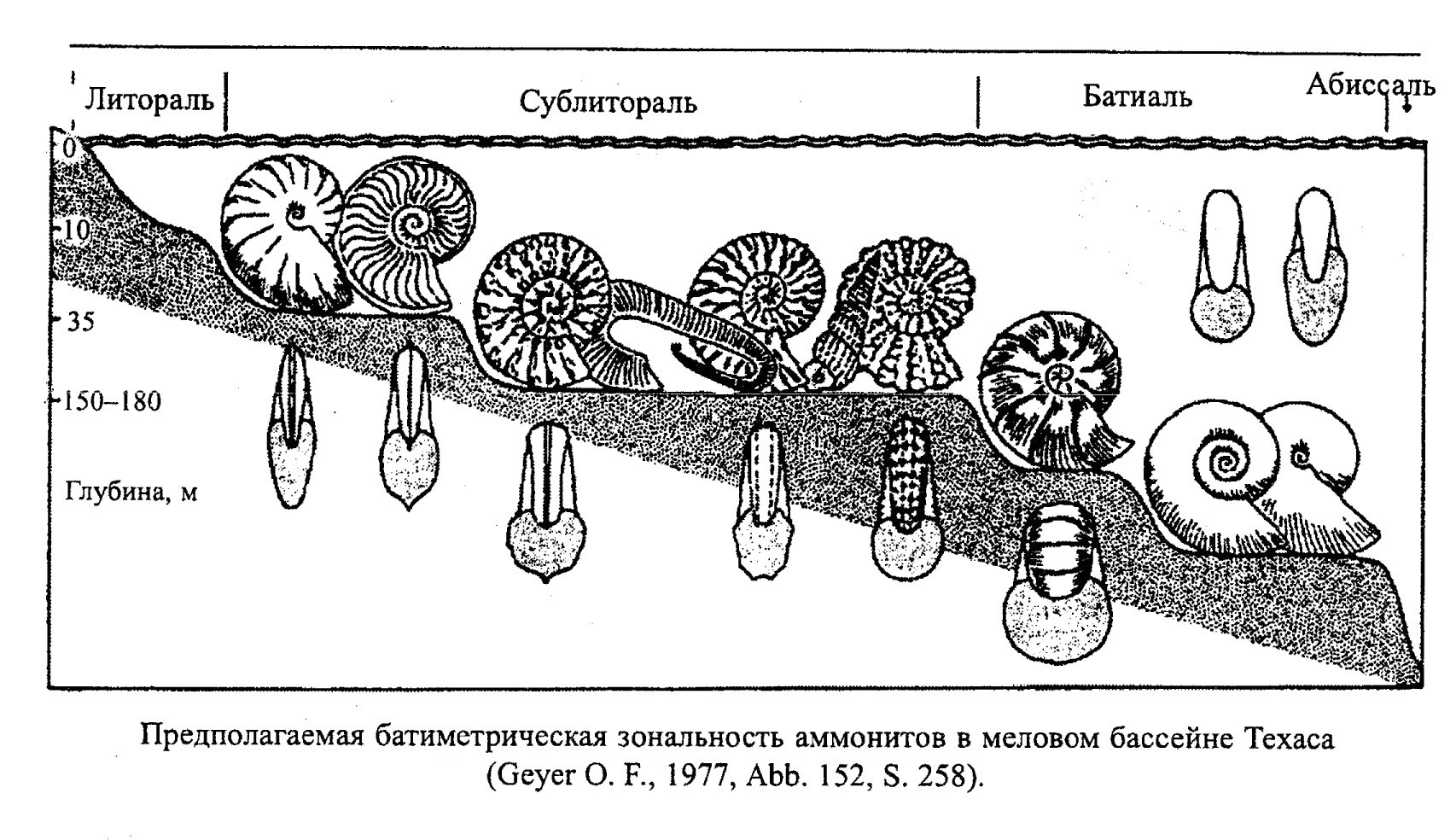
Представители подкласса Ammonoidea (рис. 40) также населяли в основном мелководные моря. Палеозойские Ammonoidea обитали на рифах, в прибрежных зарослях водорослей и обычно избегали открытых бассейнов. В мезозое эти головоногие уже широко распространены в эпиконтинентальных бассейнах и на рифах не встречаются. Аммоноидеи жили в неритовой зоне с хорошей аэрацией, не боялись активного осадконакопления, но не переносили колебаний температуры и солености воды. По мнению многих современных исследователей, аммониты были скорее всего мегапланктонными моллюсками с ограниченной способностью к активному плаванию. Были среди них и ползающие придонные животные, собирающие мелкий бентос. Девонские аммониты относились, по-видимому, к нектону. Другие палеозойские аммониты могли пассивно плавать (парить в воде) и медленно опускаться на дно. Палеозойские виды были наименее специализированными среди аммонитов. Для мезозойских аммонитов характерна гораздо более высокая специализация. Во все периоды своего существования эти аммониты были обитателями неритовой зоны, и глубина их обитания не превышала 50-200 м. Только литоцератиды и филоцератиды характерны для батиали (рис. 41). Строение раковины аммонитов позволяло им совершать медленные вертикальные миграции, возможно, суточные, что свойственно другим планктонным животным. При этом инволютные раковины выигрывали в прочности по сравнению с эволютными, поэтому аммониты с такой раковиной могли, по-видимому, опускаться на большие глубины. По способу питания мезозойские аммониты были в основном охотниками-засадчиками.
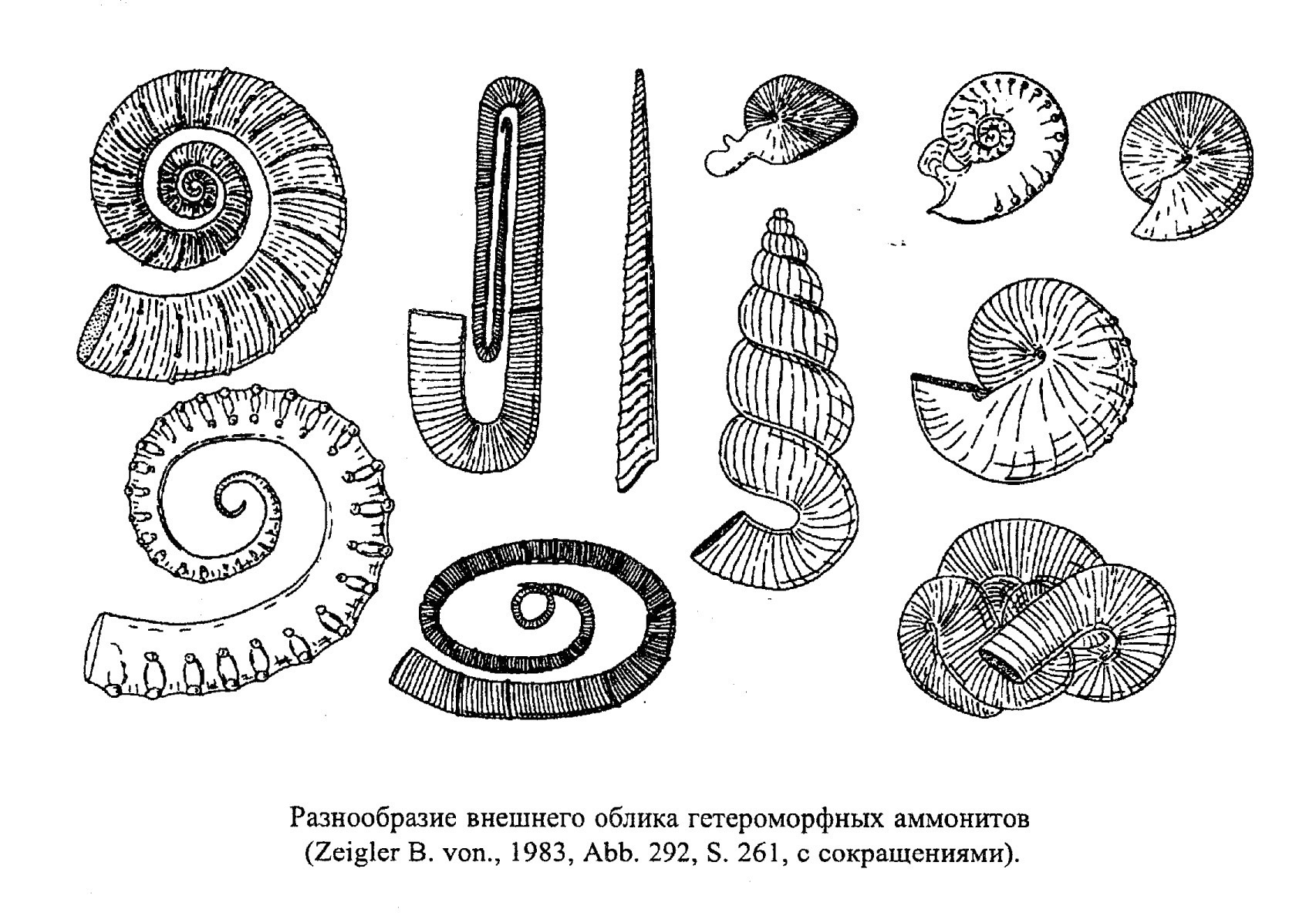
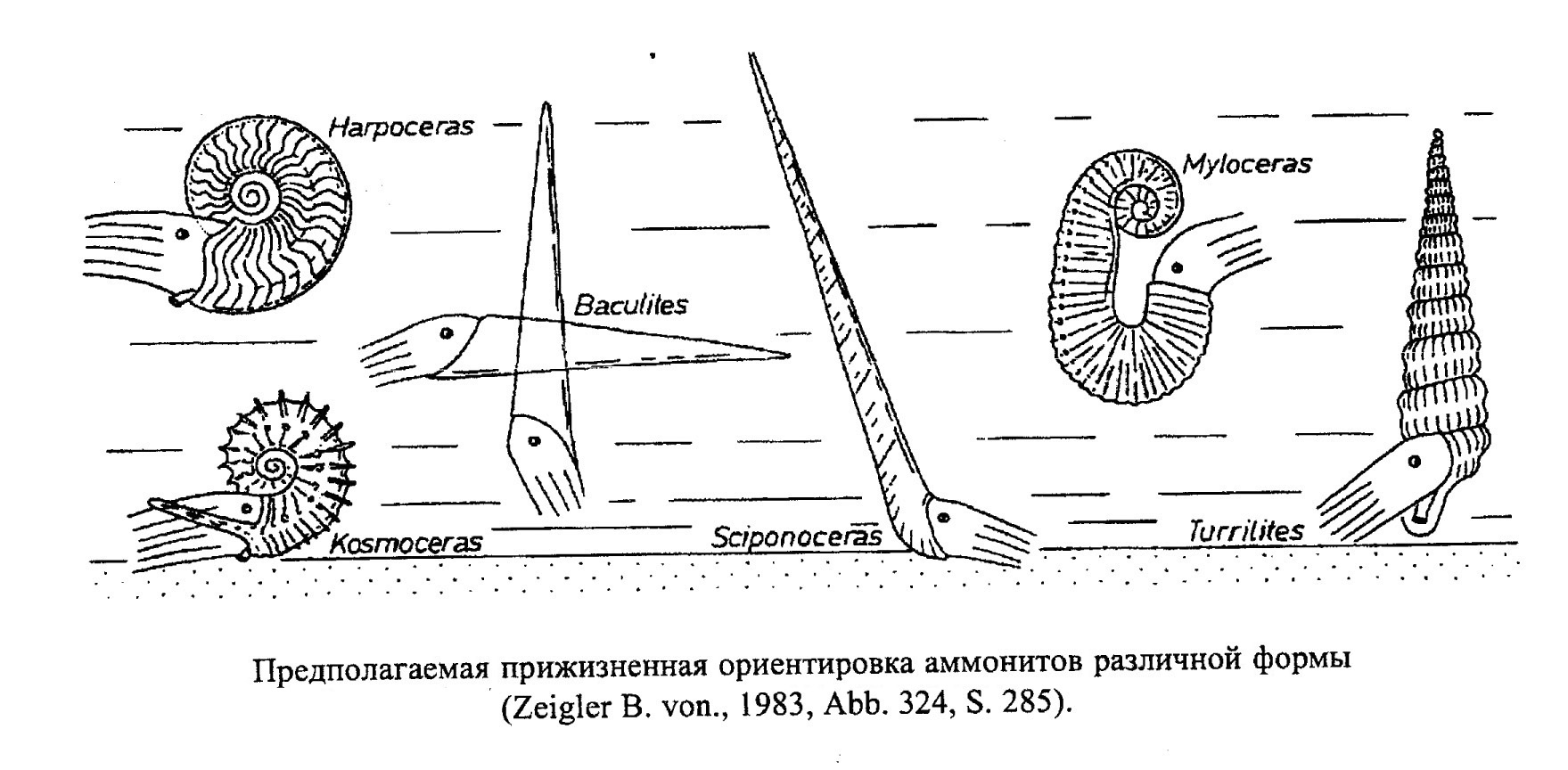
Аммониты с развитой скульптурой вели планктонный образ жизни (скульптура увеличивала поверхность раковины и делала ее более остойчивой) (обр. 385-98, 385-101). Скорее всего, так же существовали и аммониты с уплощенной суженной раковиной, которых ранее традиционно считали хорошими пловцами (их раковина также обладала большой остойчивостью). Гетероморфные аммониты, как полагают, вели планктонный образ жизни, держась среди водорослей в вертикальном положении и подстерегая добычу (рис. 42, 43).
Считают, что после смерти моллюсков раковины аммоноидей, в отличие от современных наутилусов, не всплывали и не переносились на большие расстояния, а отлагались вблизи мест их прижизненного обитания.
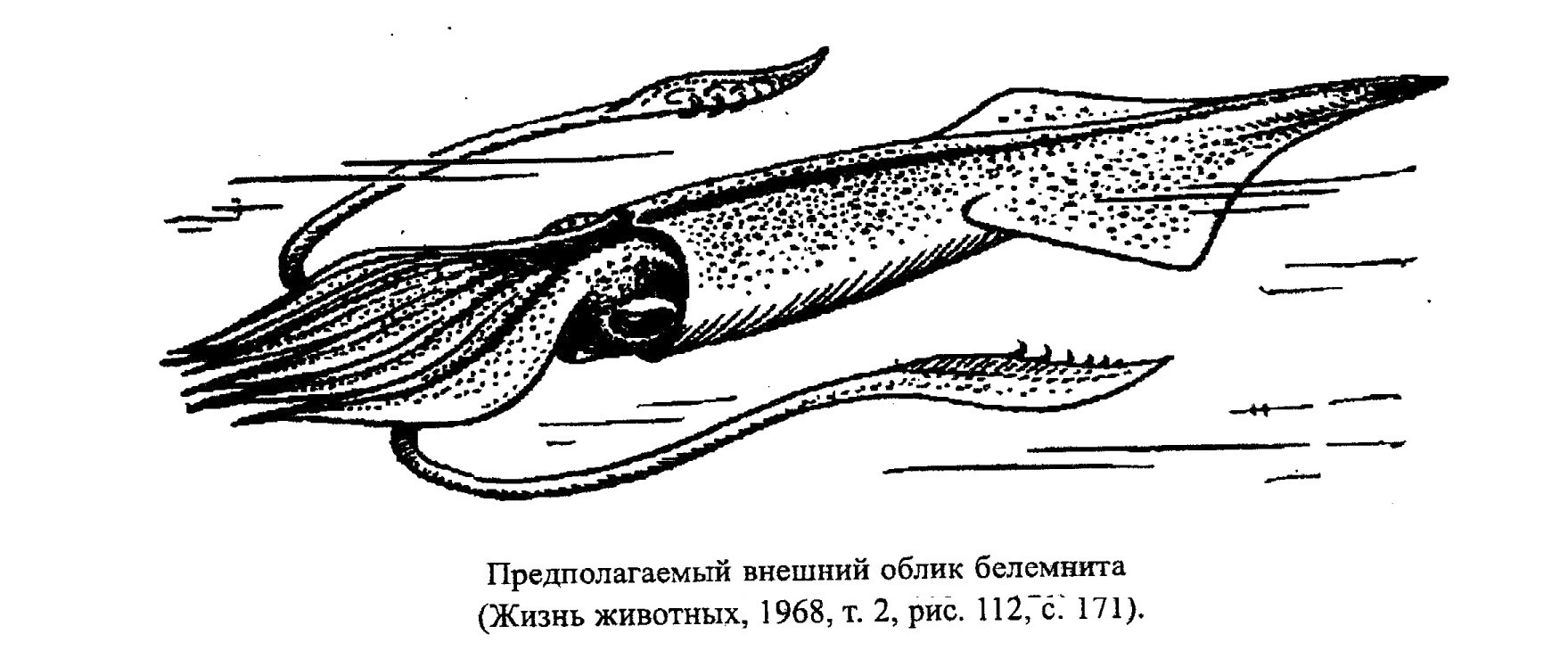
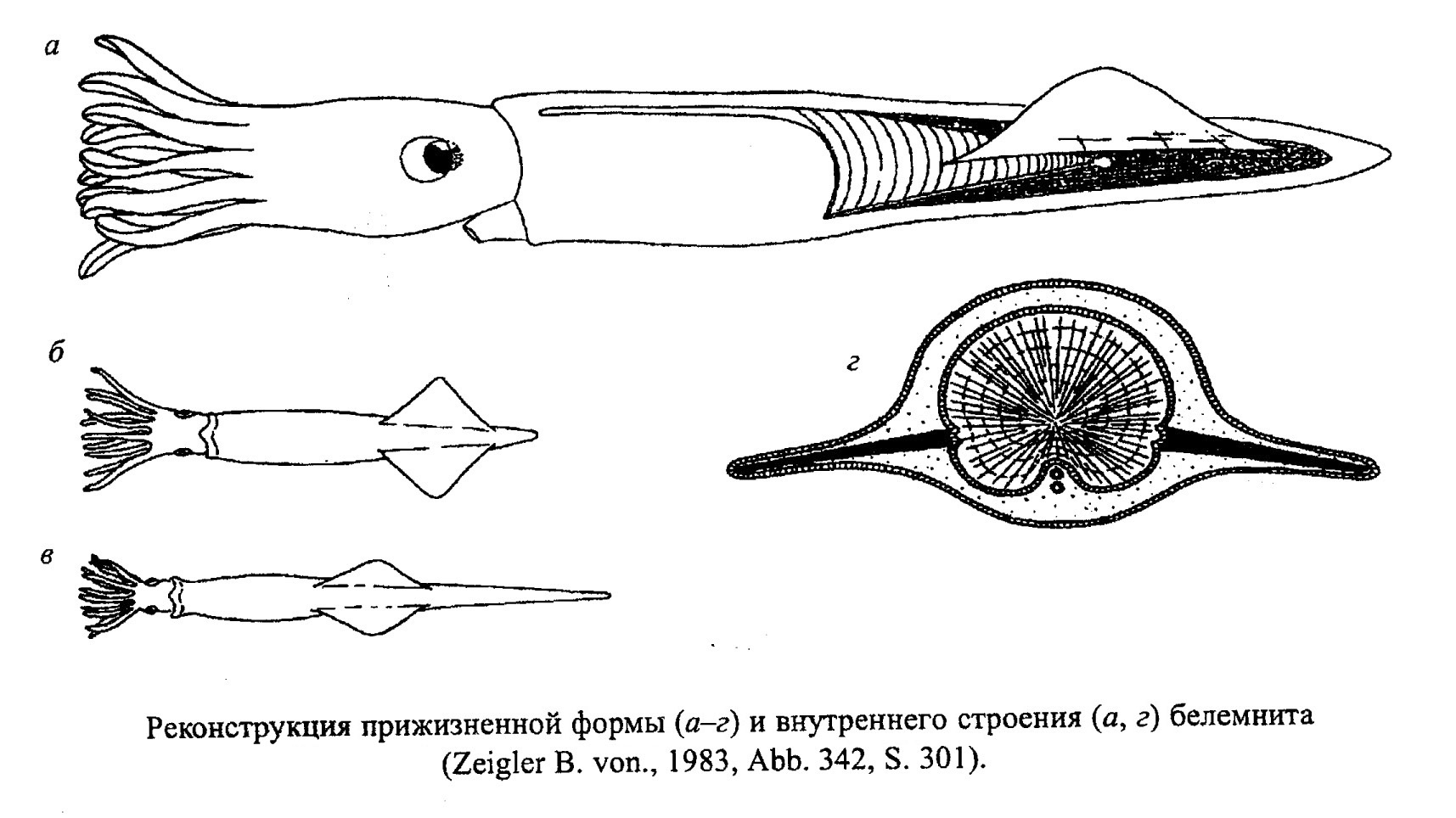
Представители надотряда Belemnoidea (подкласс Coleoidea) были распространены как в тепловодных, так и в умеренных морских бассейнах. По строению и образу жизни ближе всего к этим животным стоят современные каракатицы (сепии). Белемниты относятся к типично нектонным организмам. Судя по строению их тела, они были активными пловцами и ловкми хищниками (рис. 44, 45). Внутренняя раковина белемнитов (фрагмокон), как и наружная у аммонитов, выполняла гидростатическую функцию, при этом механическая прочность раковины не позволяла им опускаться на глубины ниже 250-300 м, поэтому белемниты населяли шельфовую зону и верхнюю батиаль.
Тип Мшанки (Bryozoa)
Мшанки - морские прикрепленные колониальные организмы-фильтраторы. Современные формы в основном обитают в мелководных бассейнах шельфовой зоны, но встречаются и глубоководные (распространены до глубин 5700 м). Многие мшанки легко переносят довольно значительные колебания солености. В солоноватоводных бассейнах мшанки биологически замешают кораллы, принимая участие в образовании органогенных построек. В отличие от кораллов эти организмы широко распространены не только в теплых, но и в холодных бассейнах, однако роль рифостроителей они могут играть только в теплых морских водах.
Мшанки - типичные обрастатели, которые селятся на твердом каменистом и скальном грунте или на гибких водорослях. Подвижные колонии мшанок легко могут стряхивать взвесь. Доминирующую роль среди известковых мшанок играют хейлостомиды. Известковые колонии мшанок имеют самую разнообразную форму (инкрустируюшие, вееровидные, грибовидные и т. д.), при этом общая форма колоний может сильно меняться в зависимости от внешних условий. Среди мшанок есть редкие сверлящие формы.
Начиная с раннего палеозоя мшанки играют роль активных породообразователей (обр. 385-111).
Тип Брахиоподы (Brachiopoda)
Брахиоподы - морские бентосные, в основном эпифаунные прикрепляющиеся или свободнолежащие сестонофаги. Небольшая группа брахиопод относится к инфаунным сестонофагам. Современные их представители живут на всех глубинах Мирового океана от зоны шельфа до 6000 м, однако ниже 300 м количество брахиопод резко сокращается и наибольшее их число приходится на глубины 100-150 м. Судя по современным представителям данной группы, большинство брахиопод - стеногалинные животные, нижней границей колебания солености для которых является 30%о. Исключение составляют представители беззамковых брахиопод рода Lingula, которые переносят резкое опреснение до 13-15%о. Остатки некоторых ископаемых палеозойских брахиопод находят в лагунных отложениях. Замковые брахиоподы мезозоя и кайнозоя обитали в бассейнах с нормальной соленостью. Одни виды современных брахиопод живут при очень низких температурах (-1,8°С в Охотском море), другие при высоких температурах (до 30°С в тропических морях). При этом большинство современных и ископаемых брахиопод все же относятся к тепловодной фауне.
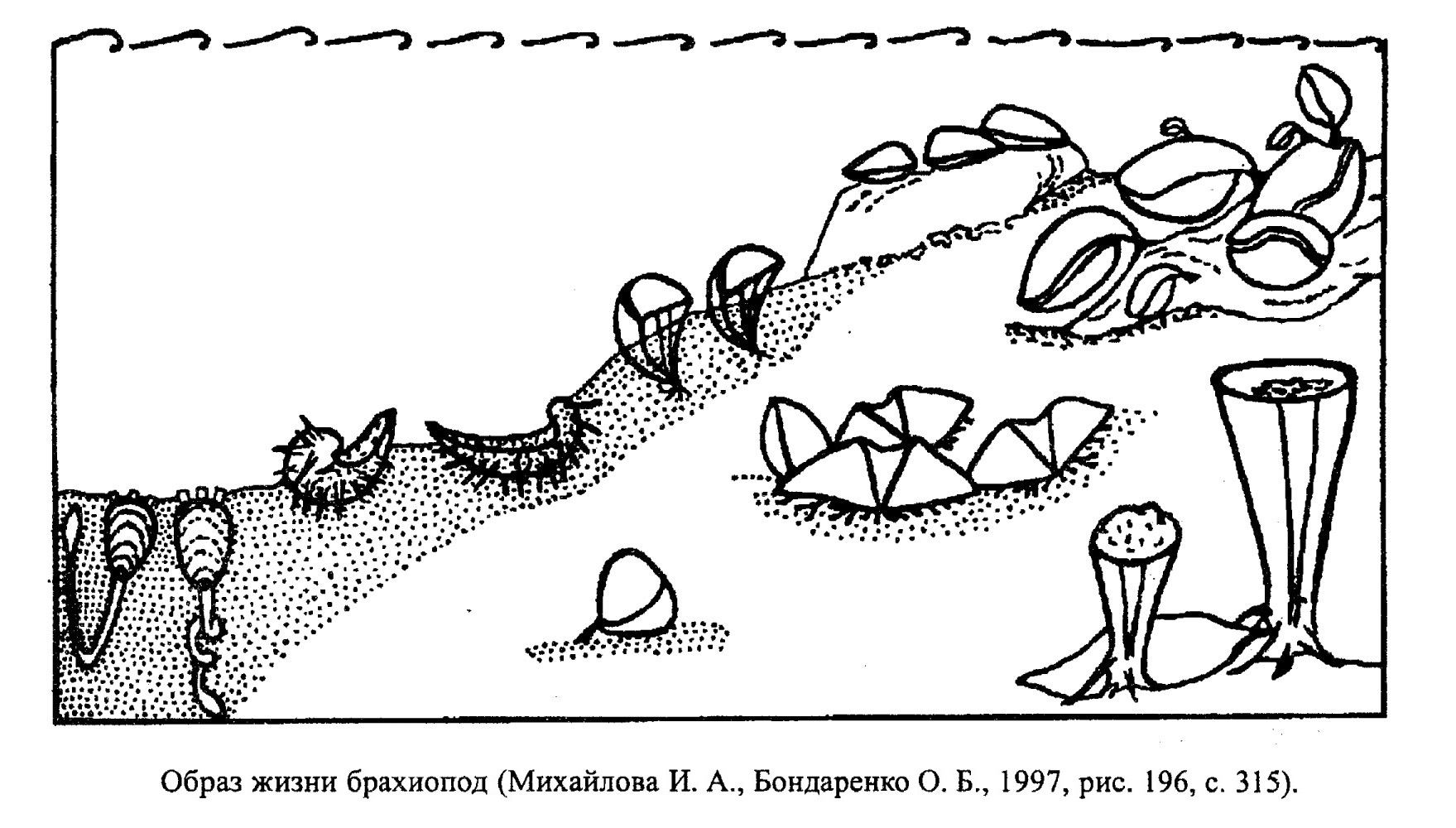
Брахиоподы сильно отличаются друг от друга по способам существования на дне бассейна (рис. 46). Среди них встречается эпифаунные (якорноприкрепленные, свободнолежащие и прирастающие) и инфаунные (зарывающиеся) формы. Первоначально (т. е. на ранних стадиях онтогенетического развития) все брахиоподы прикрепляются к субстрату ножкой, по мере роста этот образ жизни может сохраняться или сменяться другим. Поэтому среди брахиопод можно выделить несколько групп.
Первую составляют наиболее распространенные собственно якорные формы. Эти брахиоподы в течение всей жизни прикрепляются с помощью ножки. Так могли существовать только формы с очень тонкой раковиной, как, например, большинство современных и многие ископаемые (Orthida, Terebratulida, обр. 385-103а, некоторые Spiriferida и Atrypida). Якорные брахиоподы прикрепляются к твердым обломкам на дне, к вертикальному скелетному и бесскелетному бентосу, к плавающим водорослям.
Другую группу составляют формы, которые утяжелялись со временем и либо получали опору в области макушки (Rhynchonellida), либо переходили к свободному лежанию на дне. У последних часто появлялись тонкие иглы, позволяющие не погружаться в рыхлый ил (многие Productidina, обр. 385-105б), или образовывалось разрастание переднего края раковины, служившее для того же (Chonetes). Тяжелые толстостворчатые раковины с ребрами и складками свободно лежали на подвижном песчано-детритовом субстрате. Самые толстостенные (Productus) могли жить и на жестком грунте.
Якорные формы со временем могли перейти также и к прикрепленному существованию с помощью цементации. Прикреплялись они при этом либо макушкой, либо всей брюшной створкой (Crania), либо иглами к твердому дну. Как и все фильтраторы, перечисленные брахиоподы не переносят заноса рыхлым осадком. Лишь незначительное его количество они могут сбрасывать, захлопывая створки.
Последнюю группу составляют зарывающиеся беззамковые формы (Lingulida). Они живут в норках, которые роют с помощью лопатовидной ножки. Это наиболее приспособленные формы среди современных и ископаемых брахиопод, так как довольно легко переносят колебания температуры, солености воды и загрязнение бассейнов. По образу жизни к ним близки древние брахиоподы рода Obolus.
В геологическом прошлом (особенно в позднем палеозое) брахиоподы имели очень большое породообразующее значение, слагая толщи брахиоподовых известняков (обр. 385-110).
Тип Иглокожие (Echinodermata)
Подтип Кринозоа (Crinozoa)
Класс Морские лилии (Crinoidea)
Морские лилии (криноидеи) входят в состав фильтрующей донной стеногалинной фауны морских бассейнов с нормальной соленостью. Современные прикрепленные криноидеи чаще живут большими группами, образуя подводные заросли (рис. 47). Среди современных много плавающих форм с тонким изящным скелетом. Большинство современных морских лилий обитает в интервале глубин от 180 до 1000 м, но многие существуют в абиссальной зоне (до 9000 м). В мелководных зонах (в том числе и на рифах) сейчас распространены в основном бесстебельчатые криноидеи, способные активно передвигаться по дну в поисках наиболее выгодного для фильтрации положения (рис. 48). У них более грубый скелет, чем у глубоководных. Ископаемые формы с мощным скелетом также были обитателями мелководья.
Самая благоприятная температура воды для современных морских лилий - от 12 до 18°С. В таких водах наблюдается наибольшее систематическое разнообразие криноидей и максимальная их численность.
Морские лилии - активные породообразователи. В отложениях практически всего фанерозоя известны их скопления в виде криноидных известняков, сложенных как микроскопическими члениками морских лилий (в основном в зонах глубокого шельфа), так и крупными фрагментами скелетов (в мелководных зонах, где образовывались криноидные луга и биогермы) (обр. 385-125).
Подтип Астерозоа (Asterozoa)
Класс Стеллероидеи (Stelleroidea)
Подкласс Морские звезды (Asteroidea)
Морские звезды распространены в умеренных и теплых мелководных морях с нормальной соленостью. Особенно разнообразны они в тропической зоне. Большинство звезд относятся к хищникам или всеядным животным. Морские звезды активно перемещаются по дну бассейна с помощью амбулакральных ножек. Питаются они, наползая на объект добычи (кораллы, губки, двустворки, детрит, макрофиты) и обволакивая их, вывернутым наружу желудком.
Морские звезды известны с ордовика, но в ископаемом состоянии сохраняются редко (обр. 385-123).
Подкласс Офиуры (Ophiuroidea)
Офиуры - обитатели умеренных и тепловодных морей с нормальной соленостью. Они могут передвигаться по дну или зарываться в ил. Офиуры обитают на всех глубинах от литорали до абиссали. Своими гибкими щупальцами-руками офиуры захватывают взвешенные органические частицы и мелких животных из толщи воды или собирают детрит со дна бассейна, то есть относятся к группам эпифаунных и инфаунных сестонофагов и детритофагов и хищников.
В ископаемом состоянии офиуры известны с ордовика. Чаще встречаются разрозненные части скелета офиур в виде отдельных члеников. В породообразовании офиуры значительной роли не играют.
Подтип Эхинозоа (Echinizoa)
Класс Эхиноидеи, или Морские ежи (Echinoidea)
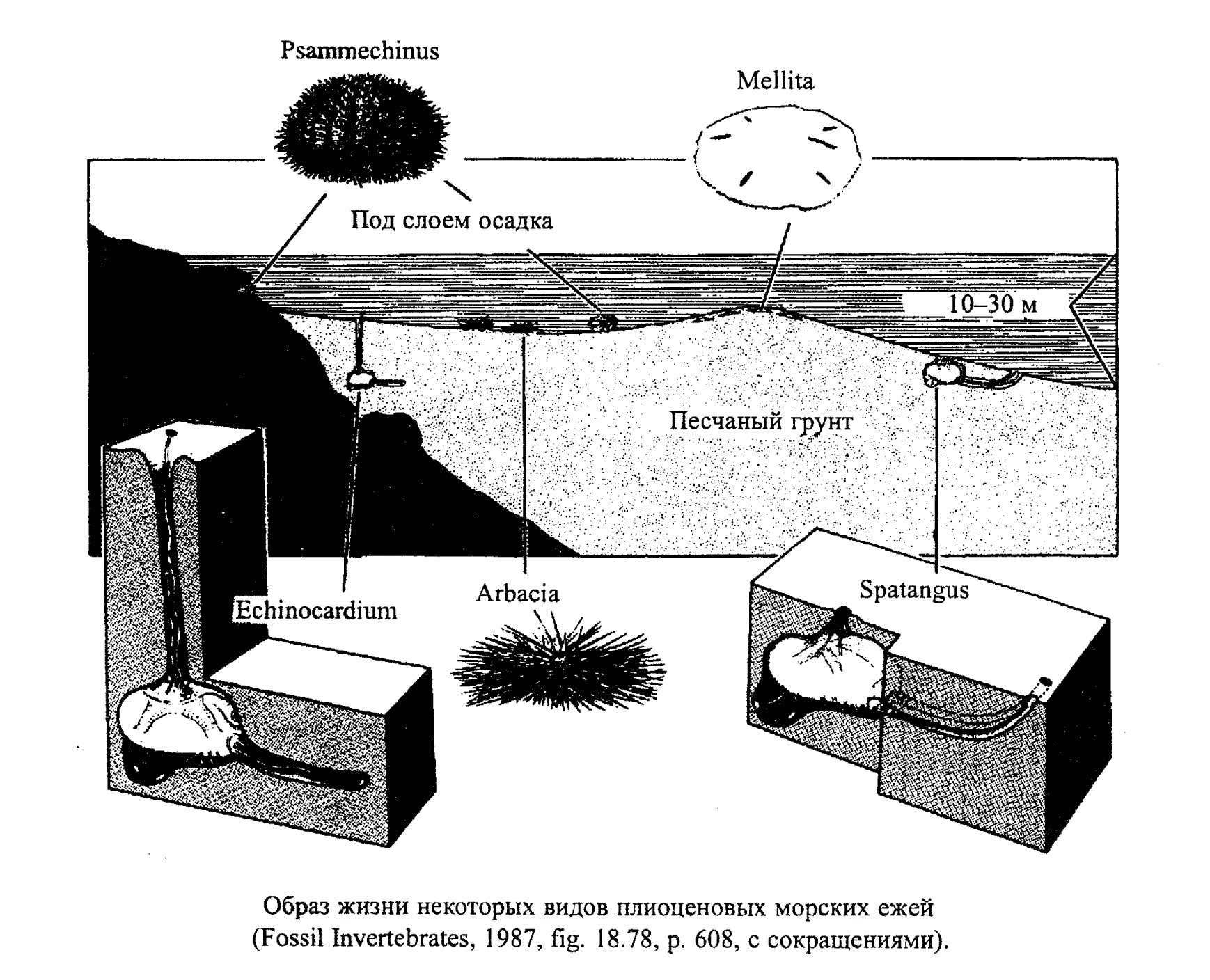
Как и все иглокожие, морские ежи - обитатели морей с нормальной соленостью. Это бентосные организмы, свободно передвигавшиеся по дну или в толще рыхлых осадков. Распространены они на всех глубинах от литорали до абиссальной зоны как в теплых, так и в холодных морях (рис. 49). В большинстве своем морские ежи всеядны. Так, современные диадемы и эхинометры питаются преимущественно водорослями, но могут поедать также и мягкие ткани склерактиниевых кораллов, и микробентос (растительноядные и хищники). Есть среди морских ежей также эпифаунные и инфаунные сестонофаги (фильтраторы) и детритофаги - сортирующие и несортирующие (илоеды).
Морские ежи различаются по способам существования на субстрате. Правильные челюстные ежи с большим числом амбулакральных ножек способны присасываться к твердому дну и выдерживать сильные волнения в береговой зоне. У таких морских ежей хорошо развитый челюстной аппарат, поэтому среди них много хищников, есть и растительноядные формы (в том числе соскребатели). Правильные морские ежи с малым числом амбулакральных ножек могут существовать лишь в относительно спокойных водах. Среди челюстных морских ежей много сверлящих форм, которые проделывают углубления в скалах и таким образом защищаются от воздействия волн. Многих ежей предохраняет от волнобоя большое число игл или прочный толстый панцирь. Такие ежи населяют прибрежные скальные зоны и рифы. В более спокойных условиях обитают правильные морские ежи с крупными иглами-ходулями (например, Cidaris, обр. 385-127), способность присасываться к субстрату у них развита слабее.
У всех неправильных морских ежей способность присасываться редуцирована. Неправильные челюстные морские ежи питаются преимущественно детритом и приспособлены к жизни на поверхности рыхлого дна и в верхних горизонтах осадка, нижняя поверхность их сильно уплощена (Clypeaster, обр. 385-130). Сердцевидные морские ежи относятся к зарывающимся детритофагам (Toxaster, Echinocorys, обр. 385-128) и играют большую роль в биотурбации осадков.
Роль морских ежей как породообразователей обычно незначительна.
Тип Полухордовые (Hemichordata)
Класс Граптолиты (Graptolithina)
Все граптолиты обитали в умеренных и теплых морях с нормальной соленостью и были фильтраторами. Среди них различают две экологические группы.
Первую (рис. 50а,б) составляют представители отряда Dendroidea (подкласс Stereostolonata) - древовидные донные прикрепленные животные, обитавшие на мелководье (например, Dictyonema, обр. 385-132). Некоторые из них могли также вести псевдопланктонный образ жизни, прикрепляясь к плавающим водорослям.
В другую (рис. 50в,г), более многочисленную группу входят представители подкласса Graptoloidea (граптолоидеи, обр. 385-131). Это пелагические животные, которые относятся к плейстонному сообществу. Колония граптолитов прикреплялась к плавательному пузырю, что позволяло этим животным существовать в приповерхностной части воды открытого шельфа. Течениями такие колонии могли разноситься на большие расстояния.
Остатки граптолитов встречаются преимущественно в аргиллитах и глинистых сланцах в виде тонких пленок из хитиноподобного вещества.
? Тип Хетогнаты (Chaetognata)
Класс Конодонты (Conodonta)
Название "конодонты" применяется для обозначения микроскопических зубовидных остатков, а также в качестве таксономического названия для группы вымерших организмов, которым эти остатки принадлежали. Последние относятся либо к типу Chaetognata беспозвоночных, либо к хордовым, сходным с миксинами и ланцетниками. По типу питания животные-конодонтоносители скорее всего, принадлежали к нектобентосным хищникам.
Современные хетогнаты относятся к пелагическим мезопланктонным животным, и распространение их имеет четкую стратификацию по глубине. Остатки видов, обитающих на больших глубинах, не встречаются в мелководных отложениях, тогда как виды, занимающие более высокие горизонты пелагиали, будут представлены и в мелководных, и в глубоководных осадках.
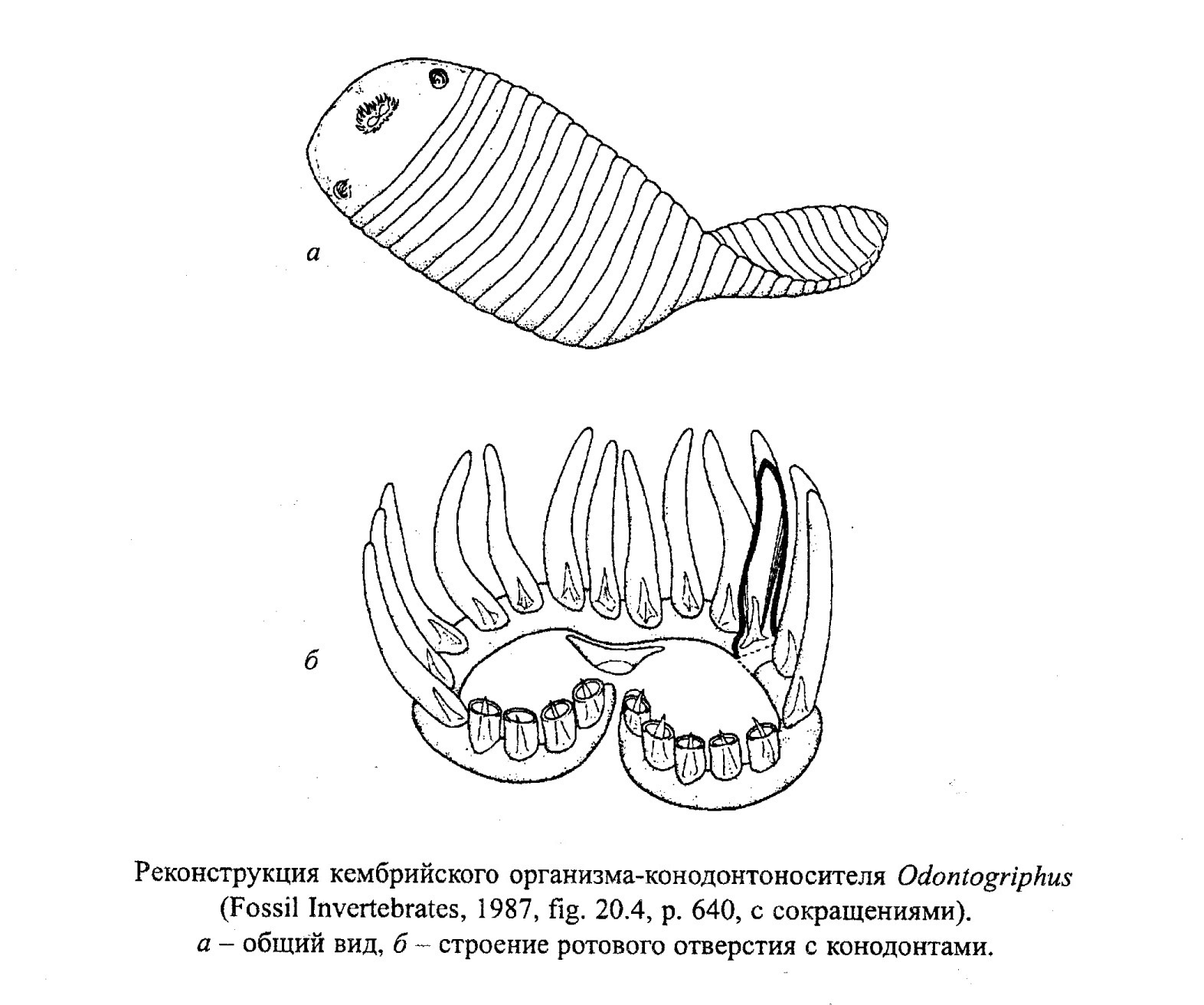
Конодонтоносители были морскими свободно плавающими неритовыми пелагическими животными (рис. 51). Это подтверждается отсутствием тесной связи их остатков с определенными литофациями. По-видимому, при этом различные группы конодонтоносителей обитали не только на разных глубинах, но и на разном удалении от берега. В настоящее время выделяют от трех до шести конодонтовых биофаций для различных периодов, которые включают
- прибрежно-морское сообщество, обитавшее в условиях изменчивой гидродинамики и перемены солености (остатки этой группы встречаются в органогенно-детритовых известняках);
- экологическую группу открытого моря, куда входят эпипелагические и мезопелагические формы (остатки этой группы особенно богаты в цефалоподовых известняках и ассоциируют с остатками рыб и морских остракод);
- рифовую группу (самую бедную).
Изменения систематического состава конодонтов в пределах пластов даже небольшой мощности позволяют улавливать очень кратковременные изменения обстановок в палеобассейнах.

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}