Экспозиция "Образ жизни некоторых современных и ископаемых организмов"
Царство Животные (Zoa, или Animalia)
Подцарство Простейшие (Protozoa)
Тип Саркодовые (Sarcodina)
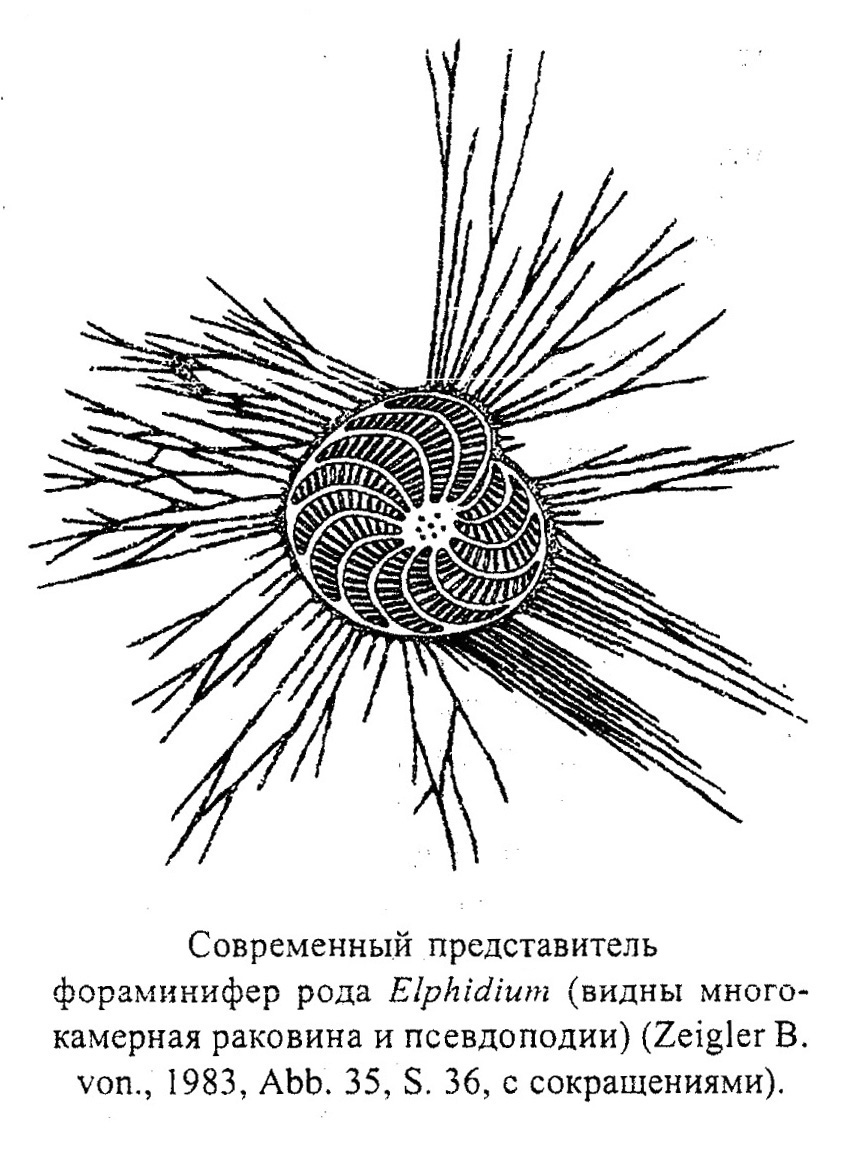
Класс Фораминиферы (Foraminifera)
Фораминиферы являются преимущественно морскими организмами (рис. 12), редкие их представители заселяют солоновато-водные и пресноводные водоемы (лагуны и эстуарии). Существуют две крупные биономические группы фораминифер: бентосные и планктонные. По способу питания среди фораминифер выделяются всеядные сестонофаги (фильтраторы), хищники и растительноядные формы.
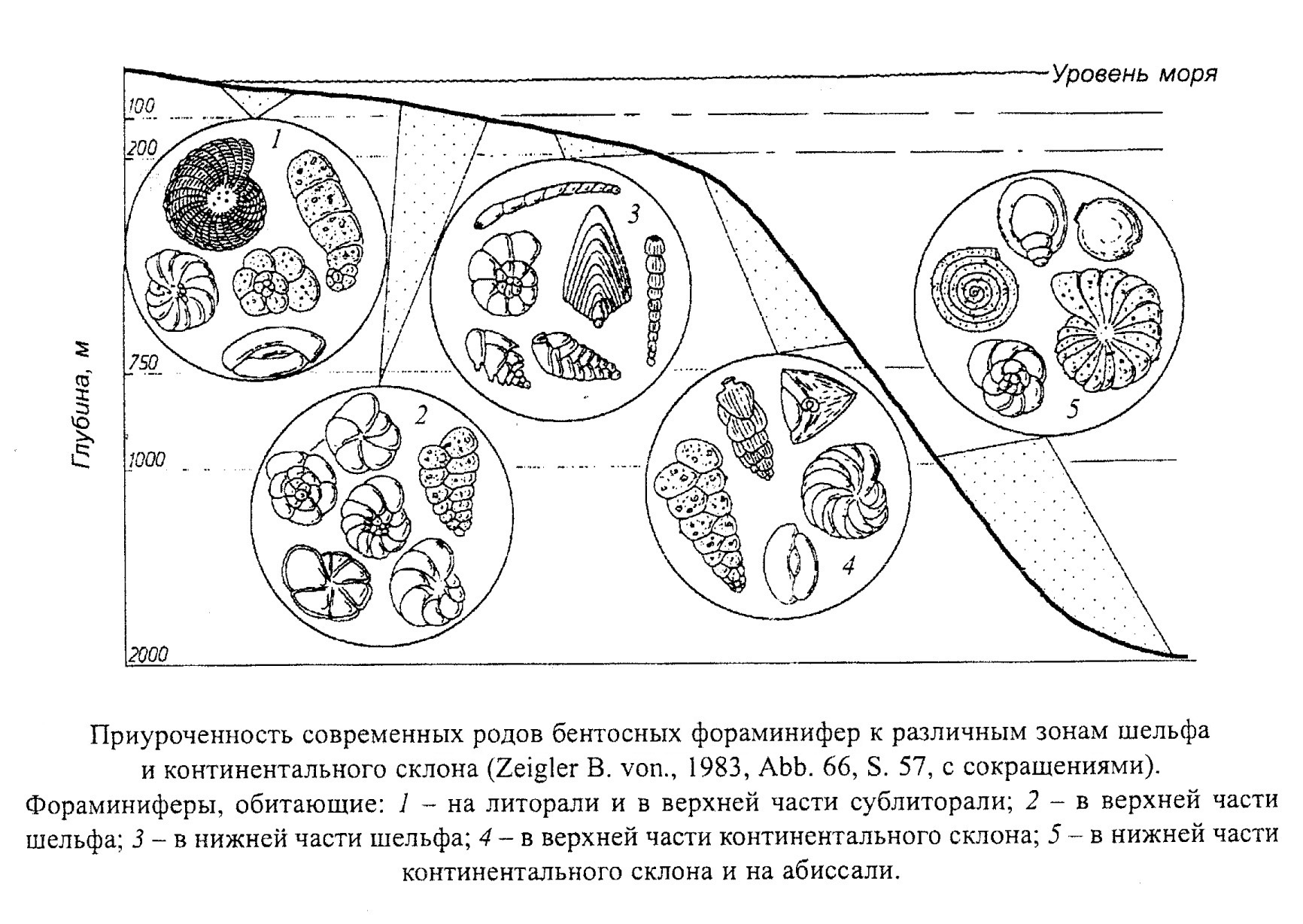
Бентосные фораминиферы наиболее многочисленны (около 3500 современных и ископаемых родов). В большинстве своем они относятся к подвижному бентосу - передвигаются по гравийному, песчаному, илистому дну или в верхнем слое осадка. Небольшая группа бентосных фораминифер ведет прикрепленный образ жизни (прикрепляются органическим или минеральным цементом). Бентосные фораминиферы заселяют преимущественно шельфовые зоны (литораль - сублитораль), но встречаются и на больших глубинах (континентальный склон - абиссаль) (рис. 13).
Планктонные фораминиферы (около 130 современных и ископаемых родов) обитают в пелагиали и только при условии нормальной солености вод.
Выделяется также группа более редких псевдопланктонных фораминифер, которые живут, прикрепляясь к плавающим предметам.
По соотношению в осадке планктонных и бентосных фораминифер судят об относительной глубине бассейна - с удалением от берега и углублением бассейна количество планктонных форм растет.
На расселение фораминифер влияет сумма факторов внешней среды (температура, соленость, глубина, химический состав воды и газовый режим грунта, прозрачность вод, их турбулентность и т. д.).
Влияние температуры воды на распространение бентосных фораминифер наиболее существенно сказывается в неритовой области, где в интервале первых 50 м она тесно связана с сезонными климатическими колебаниями. С увеличением глубины влияние климата ослабевает. Бентосные фораминиферы с секреционной известковой раковиной существуют при температурах от 2 до 36°С на глубинах до 3700-4000 м, где насыщенность вод растворенным CaCO3 составляет не менее 70%
, и раковина их не растворяется. Бентосные секреционные фораминиферы в большинстве своем являются теплолюбивыми. При этом наиболее крупные формы, такие как современные Marginopora диаметром до 3 см, ископаемые Fusulina (обр. 385-19а), Schwagerina (обр. 385-20), Orbitolina (обр. 385-23), Nummulites (обр. 385-21б), - обитатели исключительно хорошо освещенных зон мелководных тропических и субтропических морей. Гигантские размеры многих из них объясняются симбиозом с одноклеточными водорослями, благодаря которому резко возрастает скорость кальцификации их раковин.
Агглютинирующие формы приспособлены к существованию в холодных водах и на больших глубинах.
На распространение бентосных фораминифер на мелководных участках влияют сильные волнения и течения, здесь выживают главным образом прикрепленные формы и фораминиферы, живущие в толще осадка.
Современные планктонные фораминиферы по месту обитания подразделяются на три основные группы: группу тепловодного пояса (от 40° с. ш. до 40° ю. ш.) и две группы холодноводных поясов - к югу и северу от тепловодного. Большинство видов принадлежит тепловодному поясу (семейства Globigerinidae и Globorotaliidae), в холодноводных поясах распространены главным образом представители семейства Globigerinidae (рис. 14). Современные планктонные фораминиферы в основном живут на глубинах 0-300 м, причем наибольшая их концентрация фиксируется в интервале 10-50 м. Однако некоторые из них могут существовать и на глубинах до 3000 м. При этом планктонные формы могут не только "парить" в толще воды, но и перемещаться по вертикали в направлении более благоприятных условий.
Скелеты фораминифер играют большую роль в образовании осадков и пород. Они составляют значительную часть современных известковых илов, покрывающих 50% дна Мирового океана. Широко распространены фузулиновые известняки карбона и перми, орбитолиновые известняки нижнего мела, нуммулитовые известняки палеогена, основную часть которых составляют раковины крупных бентосных фораминифер.
Класс Радиолярии (Radiolaria)
Современные радиолярии обитают в океанических водах во всех климатических зонах от поверхности до абиссали (8000 м и более) в широком диапазоне температур. Это одиночные и реже колониальные организмы, которые ведут планктонный образ жизни (рис. 15). Радиолярии обитают только в акваториях с нормальной соленостью (32-36%
о). Питание у всех радиолярий гетеротрофное, они отфильтровывают и поглощают различные планктонные организмы.
В тропиках радиолярии достигают максимальных концентраций в поверхностном слое воды и на глубине 50-100 м. В Антарктике максимум их приходится на глубины около 100-300 м, а в Арктике - на глубины около 70 м и около 4000 м. Сильные волнения, переохлаждение, дожди вызывают миграцию радиолярий на большие глубины (например, от поверхности до 200 м). Обилие современных радиолярий часто связано с зонами апвеллингов, где происходит подъем биогенных элементов из придонных и глубинных горизонтов к поверхности. Однако радиолярии, особенно колониальные, довольно многочисленны и в бедных пищевыми ресурсами районах океана.
По преобладанию тех или иных видов радиолярий проводится разделение океанических вод на вертикальные зоны. Акантарии обитают от поверхности до глубин около 1000 м. Большинство из них эврибатно. Полицистины в тропиках образуют три характерные группы. Первая - поверхностная тепловодная (от 0 до 75-150 м). Вторая группа - субповерхностная, умеренно холодноводная (75-300 м). Третья группа - глубоководная (500-700 м и более). Большинство феодарий обитает в тропиках и обычно не поднимаюется выше глубин 3500 м.
Биогеографический анализ позволяет установить несколько ареалов современных радиолярий. В океанах выделяются тропические виды (широкотропические, экваториальные и тропическо-бореальные); виды умеренных областей; антарктические виды; космополитные виды (в Арктике и Антарктике существуют на глубинах не менее 200-500 м, а в тропиках на больших глубинах). Кроме того, существуют и неритические группировки, не связанные с открытым морем (среди этих радиолярий преобладают мелкие формы с мелкопористым иглистым скелетом и сильно выраженной изменчивостью). Колониальные радиолярии представляют тепловодную группу океанического поверхностного планктона.
Количество и степень сохранности радиолярий в породах зависит от физико-химических особенностей вод и осадка, в который они попадают после гибели. В теплых водах сразу после отмирания начинается растворение кремнийсодержащих скелетов, поэтому сохранность скелетов радиолярий в осадках холодноводных областей в целом лучше, чем в тропиках. При этом в мелководных бассейнах с большими скоростями осадконакопления радиолярии захораниваются быстрее и лучше сохраняются, а в глубоководных (например, в медленно накапливающихся красных глубоководных глинах) скелеты их всегда сильно корродированы. Наиболее тонкие ажурные скелеты, кроме того, обладают меньшей прочностью.
Значение современных радиолярий как породообразователей очень велико, а в предшествующие геологические эпохи было еще более высоким, о чем свидетельствует большое количество их раковин в осадочных кремнистых породах фанерозоя (фтанитах, яшмах, кремнистых сланцах, кремнях, силицилитах и др.). Встречаются радиолярии также и в терригенных и карбонатных породах (алевролиты, фосфориты, мергели, известняки, доломиты и т. д.). Породы с радиоляриями известны в отложениях всех систем фанерозоя.
Подцарство Многоклеточные (Metazoa)
Надраздел Примитивные многоклеточные (Parazoa)
Тип Губковые (Spongiata)
Класс Губки (Spongia)
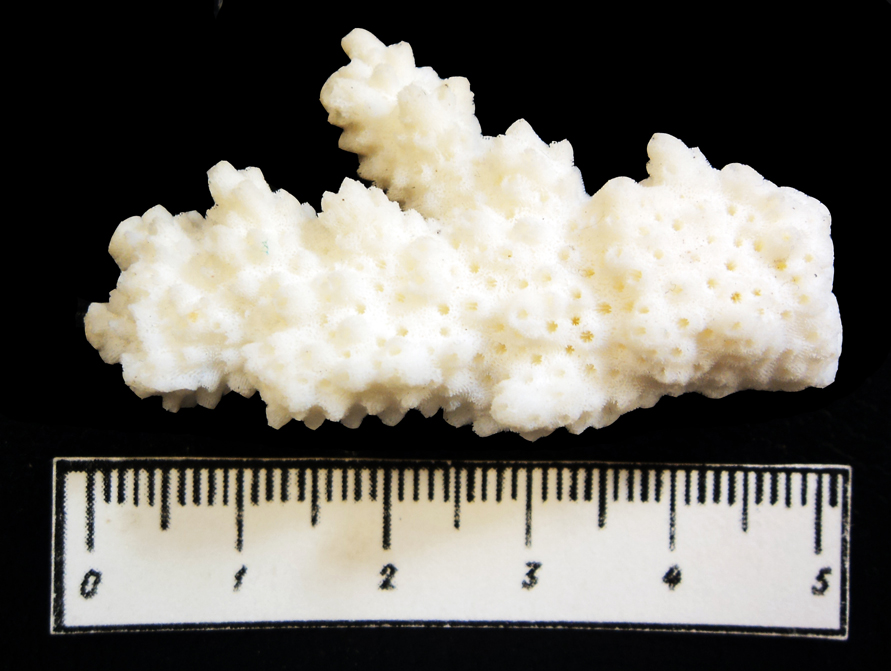
Большинство губок - обитатели морей с нормальной соленостью. Лишь немногие виды распространены в пресноводных водоемах. Живут губки как в теплых, так и в холодных водах, однако в тепловодных бассейнах их систематическое разнообразие гораздо выше. Губки - типичные сестонофаги, они относятся к сидячему бентосу и предпочитают твердый субстрат, на котором образуют крупные поселения. На илистом дне губки не выживают. Глубины обитания этих организмов - от приливно-отливной зоны до абиссали, при этом максимальной численности они достигают на шельфе. Известковые губки обычно распространены на мелководье, кремнистые - в глубоководных зонах. Как и для всех фильтраторов, для губок большое значение имеет подвижность воды, с которой связан привнос пищевых частиц. Внешняя форма губок одних и тех же видов может сильно меняться в зависимости от внешних условий (характера дна, силы и направления течений, глубины и других факторов) (обр. 385-25а, 385-25б, 385-29).
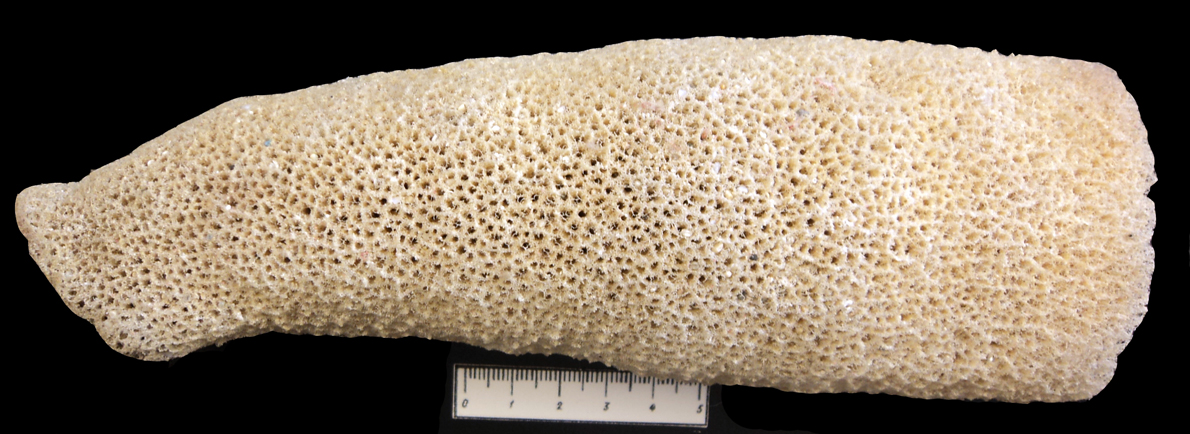
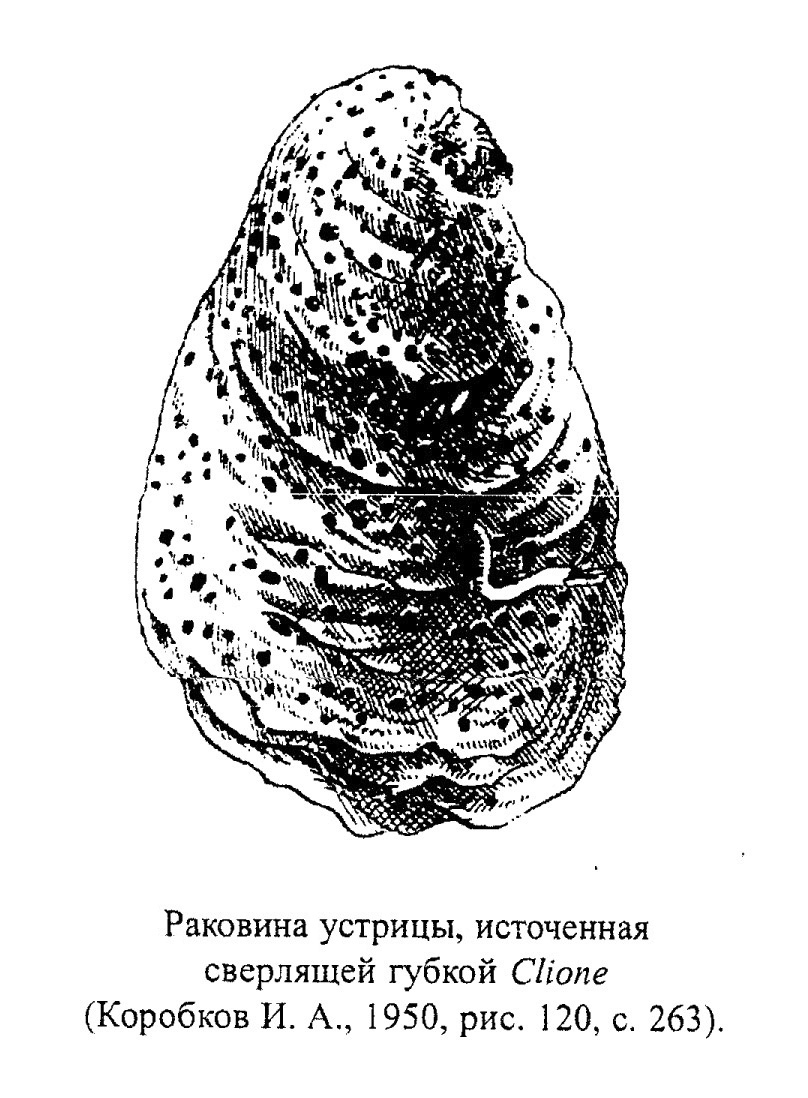
Особую группу составляют сверлящие губки (например, современные и ископаемые представители семейства Clyonidae). Клионы проделывают в твердом субстрате (раковины живых и мертвых моллюсков, колонии кораллов и т. д.) ходы диаметром 1-5 мм и поселяются в них, оставляя на поверхности небольшие выросты тела. Сверлят губки с помощью специальных "скребущих" клеток, выделяющих кислоту. Сами губки Clyonidae мягкие и в ископаемом состоянии не сохраняются (рис. 16). Сверлящие губки разрушают раковины устриц, колонии кораллов, разрыхляют скальные породы (рис. 17) (обр. 385-24). Глубина обитания современных клион не превышает 100 м.
Известковые и кремнистые губки являются важными породообразователями. Первые из них принимали участие в формировании рифов (рис. 18), вторые - в образовании спонголитов (кремнистых пород с рассеянными спикулами).
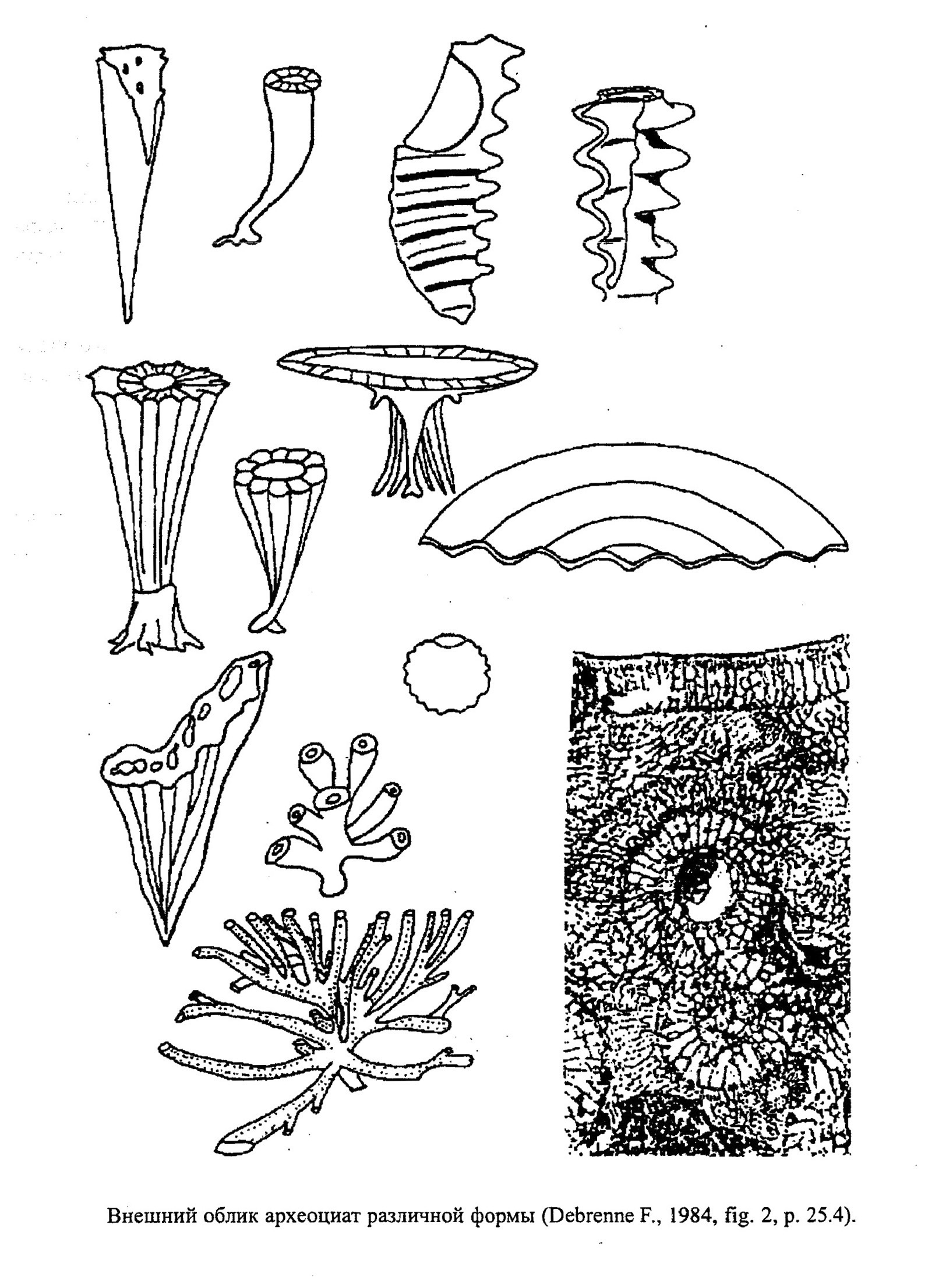
Тип Археоциаты (Archaeocyathi)
Археоциаты - фильтраторы, относящиеся к прикрепленному бентосу, представленные одиночными и колониальными формами (рис. 19). Они обитали в тепловодных бассейнах нормальной солености. Глубины их существования, судя по вмещающим осадкам, обычно составляли 5-20 м (реже до 50 м). Археоциаты предпочитали твердое каменистое или скальное дно бассейнов со слабой и средней динамикой вод. Внешняя форма представителей одних и тех же видов была весьма изменчивой в зависимости от внешних условий. Археоциаты - одни из первых рифостроителей Земли, в раннем кембрии они образовывали биогермы и биостромы различной величины и морфологии (обр. 385-31).
Надраздел Настоящие многоклеточные (Eumetazoa)
Раздел Радиально-симметричные (Radiata)
Тип Стрекающие (Cnidaria)
Среди ископаемых и современных книдарий (стрекающих) много групп, сыгравших огромную роль в процессах осадконакопления. Многие из них входят в число главнейших рифообразующих организмов фанерозоя. Это строматопораты, хететиды, гелиолитоидеи, табуляты, ругозы, шестилучевые кораллы (склерактинии), в меньшей степени - восьмилучевые кораллы. В настоящее время основными рифостроителями являются склерактинии.
Судя по морфологии скелета ископаемых кишечнополостных других групп (строматопораты, гелиолитоидеи, табуляты, ругозы), условия их существования были очень сходными с условиями жизни современных склерактиний, о которых будет сказано далее. Все они принимали активное участие в образовании древних рифов.
Класс Коралловые полипы (Anthozoa)
Склерактинии (шестилучевые кораллы с твердым известковым скелетом) являются обитателями морей с нормальной соленостью. По типу питания это сестонофаги и хищники, для многих склерактиний большую роль в питании играет также симбиоз с одноклеточными водорослями. Все они относятся к прикрепленному бентосу. Склерактинии делятся на две крупные биономические группы - герматипные (колониальные рифостроящие, обр. 385-39, 385-41, 385-45) и агерматипные (одиночные и колониальные кораллы, не образующие органогенных построек, обр. 385-37).
Главным условием существования герматипных кораллов является достаточная освещенность, поскольку основная их особенность - симбиотические отношения с одноклеточными водорослями зооксантеллами. Эти водоросли внедряются в мягкие ткани кораллов и осуществляют процесс фотосинтеза, поглощая углекислый газ, выделяемый кораллами, и поставляя им кислород и другие продукты обмена веществ. В результате этих взаимовыгодных отношений кальцификация скелета герматипных склерактиний резко усиливается, что ведет к возрастанию скорости роста колоний. Герматипные кораллы обитают в тропических и субтропических морях со среднегодовыми температурами от 18 до 40°С, однако наиболее благоприятной для их существования является температура 26°С. Глубина их обитания колеблется в пределах 0-20 м (реже до 60 м).
Рост агерматипных кораллов не зависит от процессов фотосинтеза, поэтому они распространены до глубин 6000 м и могут существовать при температурах всего в несколько градусов.
Для жизни большинства кораллов наиболее благоприятны прозрачные бассейны с подвижными водами. В мутной воде кораллы сбрасывают оседающую взвесь с помощью движения щупальцев. Кроме того, быстрорастущие герматипные формы при заиливании могут образовывать высокие кустистые колонии, что предохраняет их от заноса осадками. Однако сильного заноса илом не выносят ни герматипные, ни агерматипные кораллы.
Под влиянием различных внешних факторов (движение воды, занос илом, освещенность, периодические осушения и т. д.) внешняя форма кораллов, принадлежащих к одному и тому же виду, может очень сильно меняться (от уплощенной инкрустирующей до сферической и ветвистой) (рис. 20).
Раздел Двусторонне-симметричные, или Трехслойные
(Bilateria, или Triblastica)
Подраздел Первичноротые (Protostomia)
Тип Кольчатые черви (Annelides)
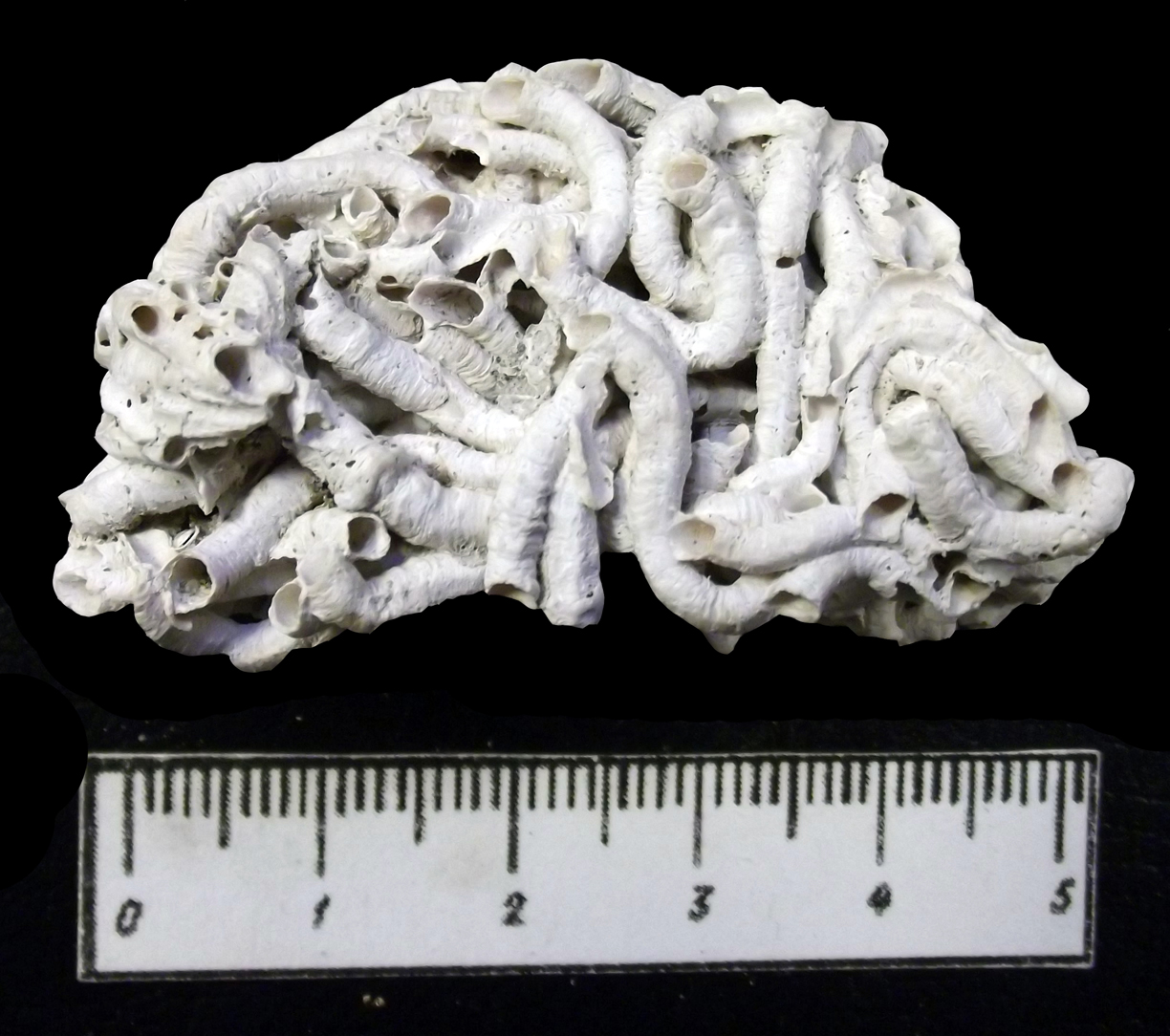
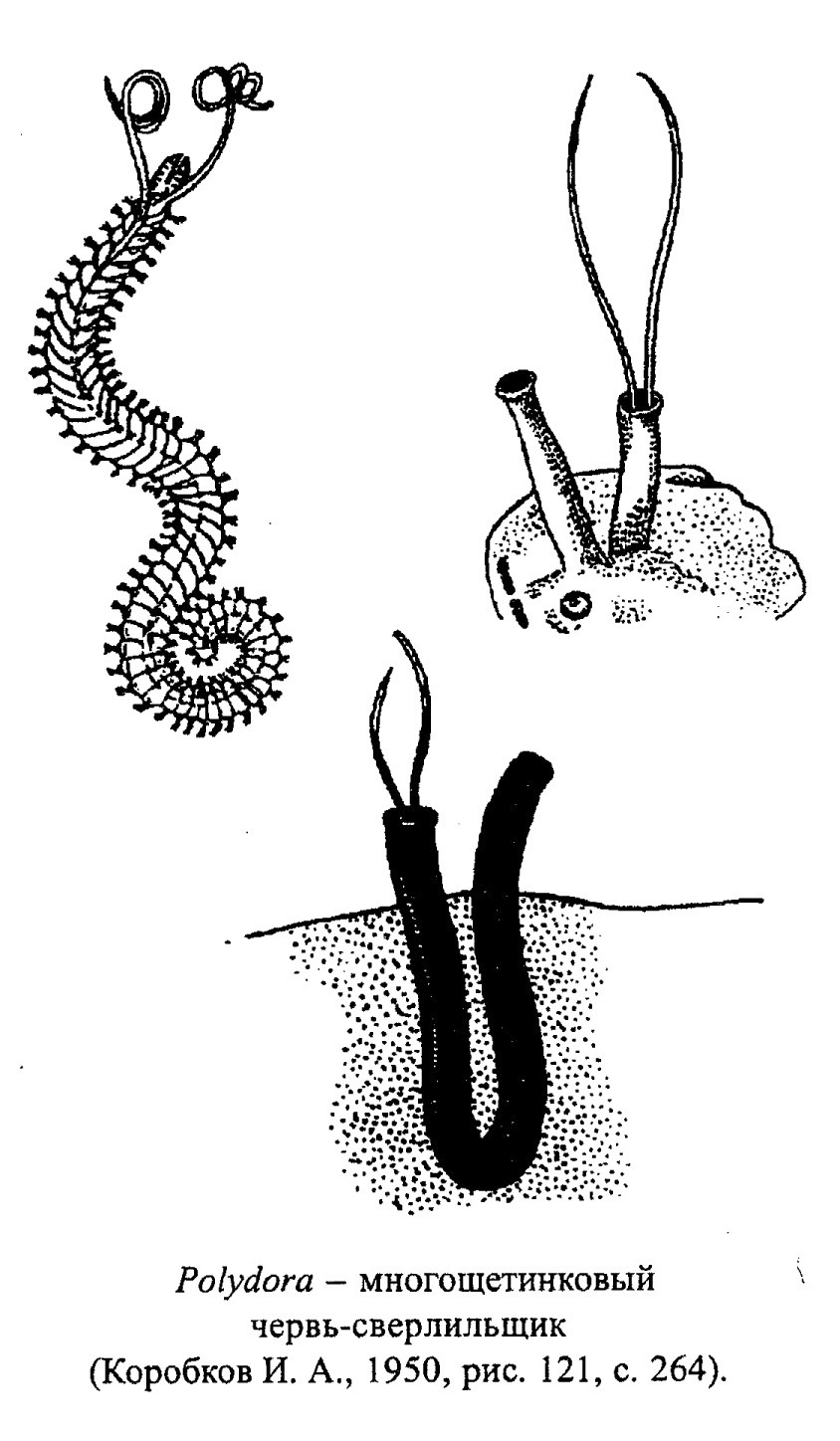
Морские кольчатые черви обитают в теплых и умеренных водах. Среди них выделяют две группы. К первой принадлежат черви, не имеющие скелета, - это илоеды, соскребатели и хищники. В ископаемом состоянии от них остаются лишь остатки челюстного аппарата (сколекодонты) и многочисленные следы перемещения в иле (фукоиды). Вторую группу составляют фильтраторы, относящиеся к сидячему бентосу, - это представители семейства Serpulidae (обр. 385-114). Мелкие серпулиды (например, Spirorbis) прикрепляются путем цементации к неподвижным предметам на дне или к раковинам живых организмов и выделяют защитную трубку (обр. 385-117). Другие серпулиды строят прочные извилистые известковые трубки на скальных поверхностях, часто покрывая их сплошным слоем, формируя довольно крупные органогенные постройки - серпулиты. В основном это формы, обитающие в теплых морских мелководных бассейнах с прозрачной подвижной водой, часто принимающие участие в процессе образования рифов. Среди червей-сестонофагов есть и сверлильщики (черви Polydora) (рис. 21).
Породообразующая роль серпулид, формирующих органогенные постройки, очень велика.
Тип Членистоногие (Arthropoda)
Подтип Трилобитообразные (Trilobitomorpha)
Класс Трилобиты (Trilobita)
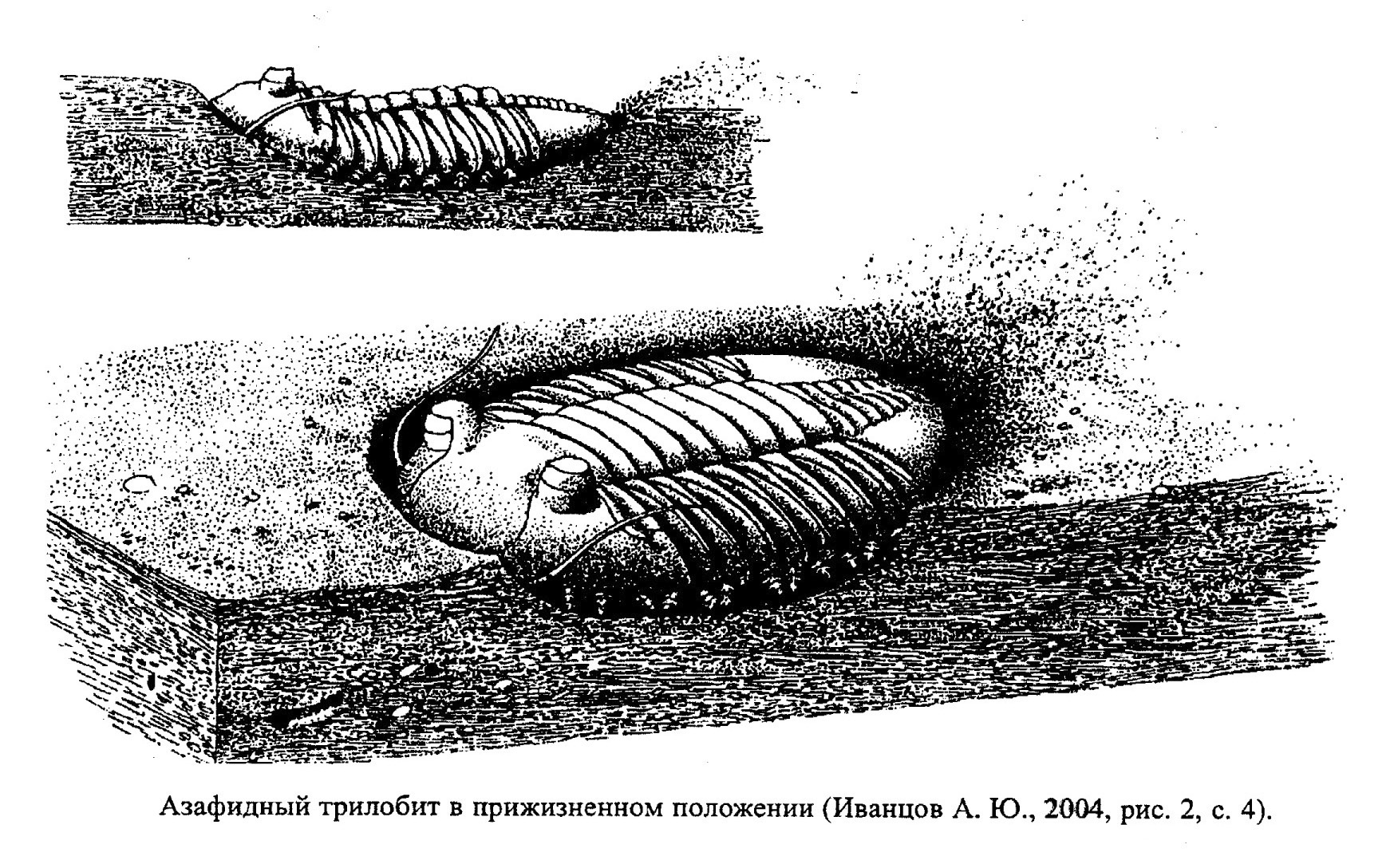
Трилобиты были обитателями умеренных и теплых мелководных морей с нормальной соленостью. Они принадлежали преимущественно к донным животным, которые обычно ползали, хотя многие из них и обладали некоторой плавучестью (рис. 22а, 23). Существовали также планктонные и нектонные формы. Возможно, некоторые трилобиты, как многие современные ракообразные, жили, прикрепляясь к плавучим водорослям.
Челюстной аппарат трилобитов был слабо развит, поэтому питались они, скорее всего, сестоном, органическими остатками и мелким детритом или донной растениями. Возможно, были среди них и хищники. Обитатели мягких грунтов зарывались в ил, а твердых - обитали на поверхности дна.
На распространение трилобитов более всего влияла соленость бассейна и тип грунта. Некоторые из них переносили опреснение и осолонение. На твердом субстрате жили наиболее активные формы с хорошо развитой мускулатурой, о чем свидетельствует расчлененный панцирь. Трилобиты, жившие в зонах активной гидродинамики, имели прочный толстостенный панцирь. На рыхлых грунтах обитали трилобиты с широким, слабо выпуклым панцирем. Для них характерны выросты, шипы и широкий лимб вокруг головного щита, расположенные в одной плоскости, что позволяло им не погружаться в рыхлый осадок (обр. 385-119). У зарывающихся трилобитов часто редуцированы органы зрения. У некоторых зарывающихся форм глаза располагались на особых выростах (стебельках). У тех трилобитов, которые вели нектонный образ жизни, глаза располагались у края головного щита, что, по-видимому, позволяло увеличить обзор. Многие трилобиты имели способность сворачиваться, защищаясь от хищников (обр. 385-121).
В отложениях нижнего палеозоя остатки трилобитов часто образовывали крупные скопления (обр. 385-120).
Подтип Ракообразные (Crustacea)
Класс Ракушковые ракообразные, или Остракоды (Ostracoda)
Остракоды - водные организмы, приспособившиеся к обитанию в морских, солоноватоводных и пресноводных водоемах разных широт (рис. 22б). Некоторые группы эвригалинны и способны обитать в гиперсоленых водах с концентрацией солей до 200%
о. Другие обитают во влажной листве, в почве лесов и даже в горячих источниках и глубоких колодцах пустынь. Подавляющее большинство остракод являются свободноживущими бентосными формами. Меньшую по численности группу составляют пелагические остракоды, способные к длительному плаванию (встречаются на глубинах до 4000 м). По типу питания большинство остракод всеядно, но есть среди них фильтраторы, растительноядные, трупоядные формы и детритофаги.
На морском шельфе, особенно в прибрежной зоне, где существуют сложные системы мелководных лагун и эстуариев, остракоды образуют значительное число специфических сообществ, связанных с определенными глубинами, грунтами, гидродинамикой и соленостью. Есть формы, адаптированные к существованию в холодных и богатых кислородом глубоких придонных водах (психосферические остракоды). По комплексам остракод можно восстанавливать различные обстановки осадконакопления (мелководный шельф, гемипелагическая обстановка, глубокий холодноводный шельф). Остракоды, населяющие различные биотопы, чрезвычайно важны для палеогеографических построений и особенно для сопоставления морских и континентальных отложений.
В нормальноморских отложениях раковины остракод редко встречаются в больших количествах и, как правило, не являются породообразователями. Но в солоноватоводных или краевых морских бассейнах с соленостью, отклоняющейся от нормальной, некоторые группы остракод получают чрезвычайно пышное развитие и накапливаются в большом количестве, образуя остракодовые известняки.
Тип Моллюски (Mollusca)
Класс Брюхоногие моллюски (Gastropoda)
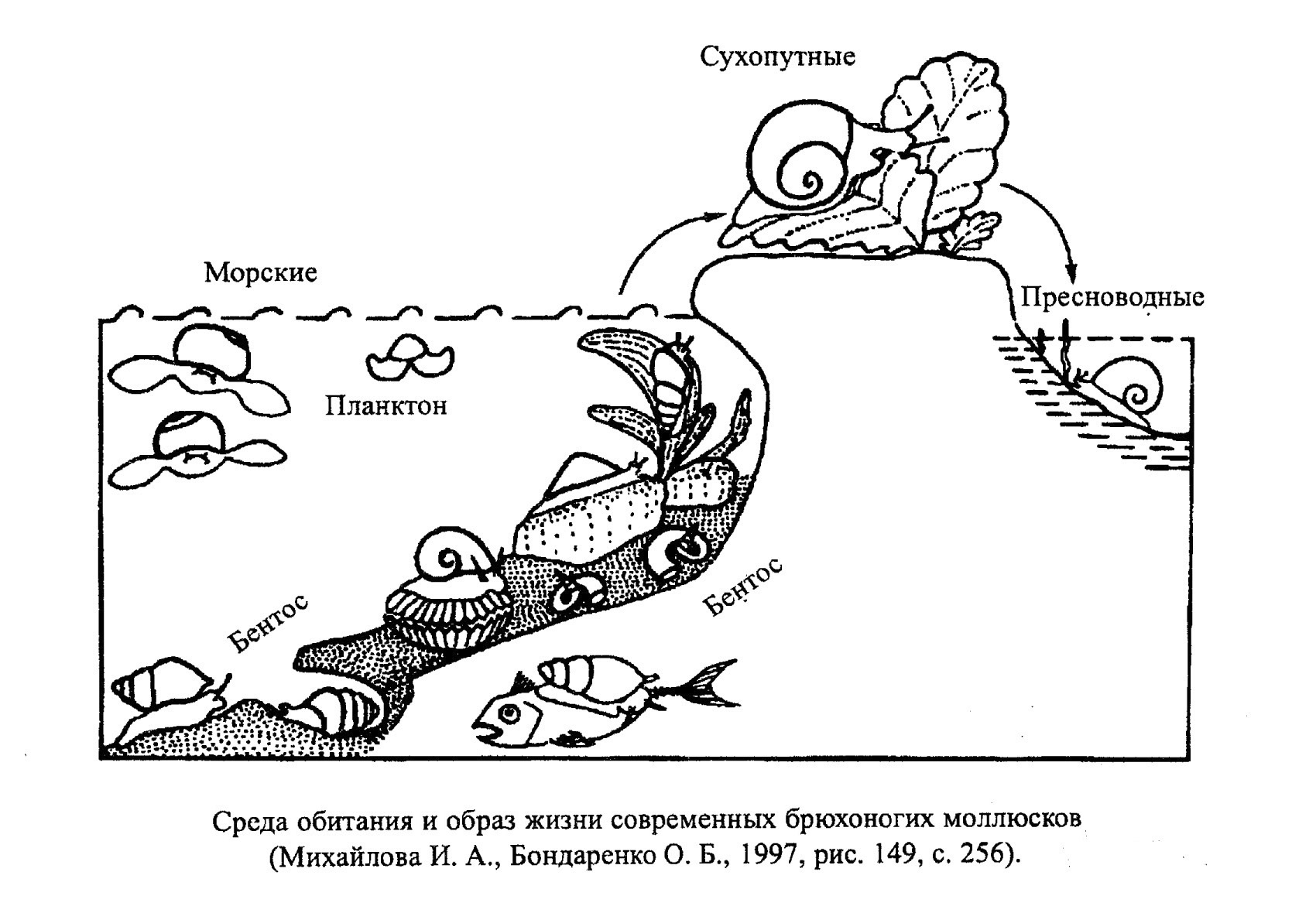
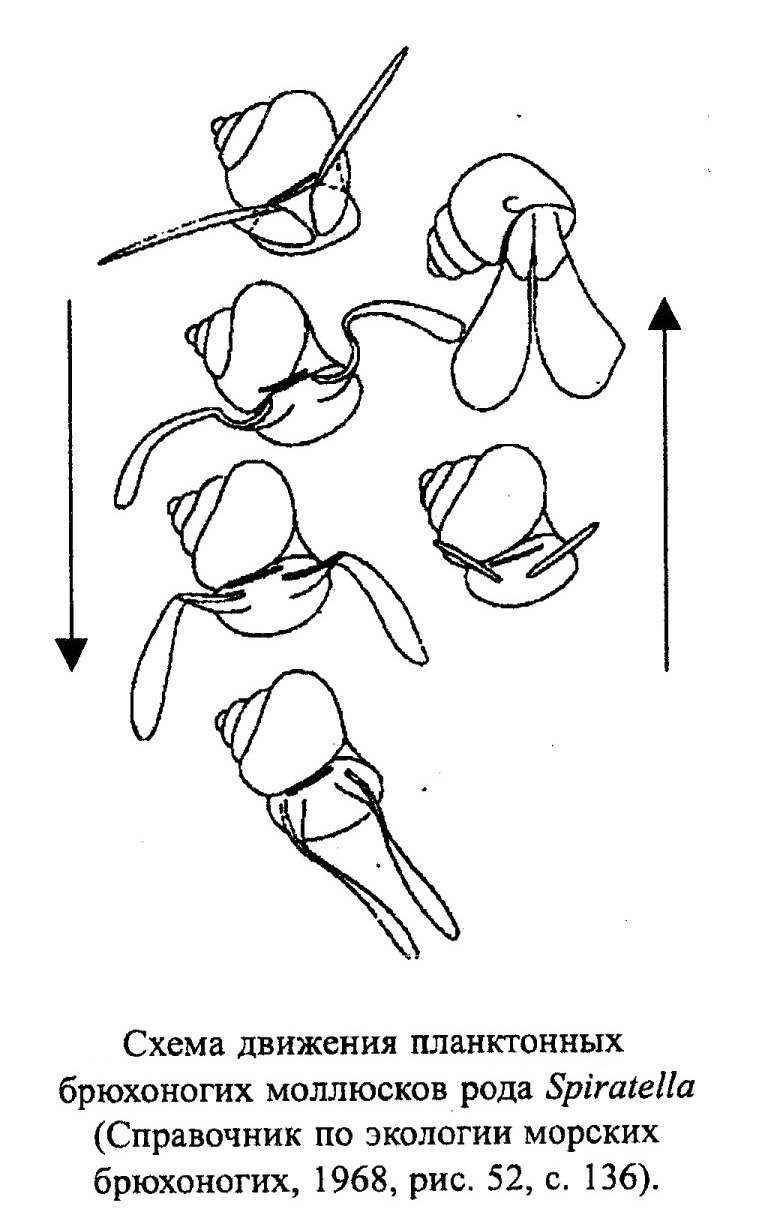
Подавляющее большинство брюхоногих моллюсков (гастропод) обитает в морях нормальной или близкой к нормальной солености, населяя прибрежные мелководные участки дна, хотя есть бентосные виды, опускающиеся до абиссали (рис. 24). Небольшая группа морских гастропод (птероподы) ведет планктонный образ жизни (рис. 25). Меньшая часть гастропод населяет пресноводные водоемы или обитает на суше. Большинство брюхоногих моллюсков относится к активным животным.
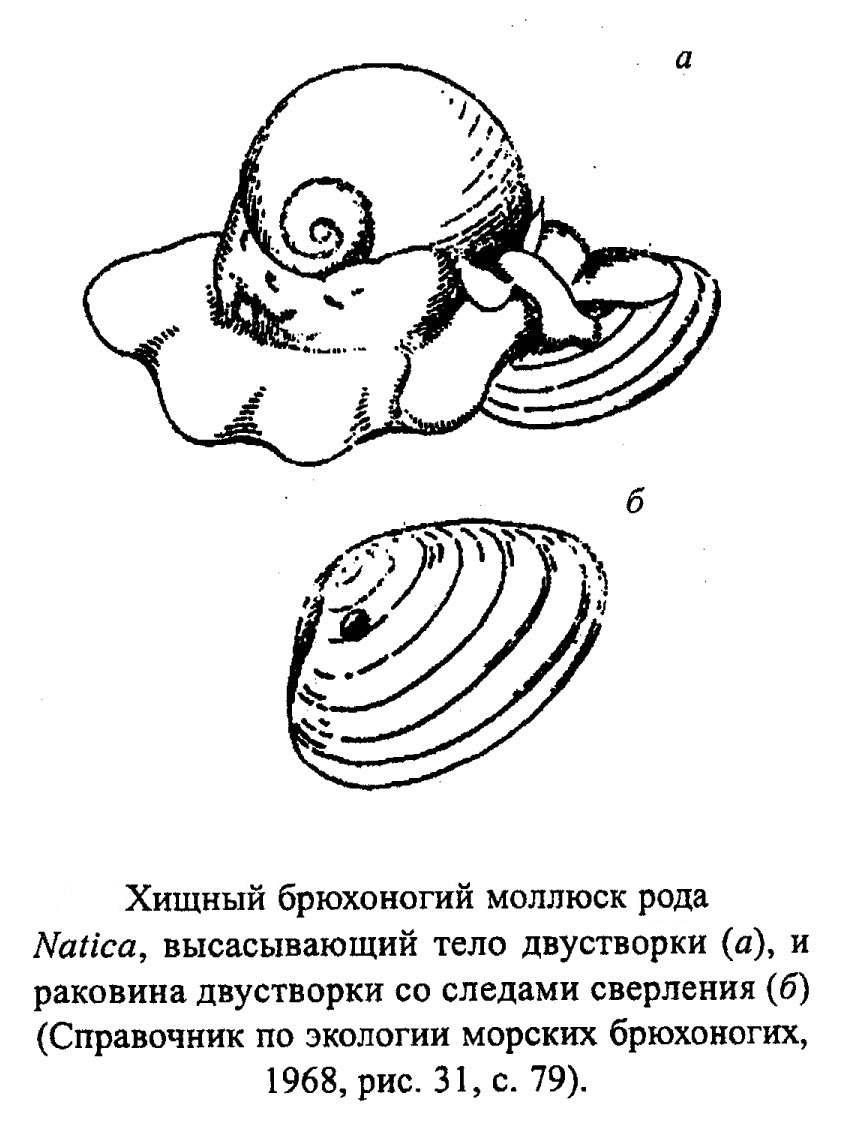
У гастропод можно встретить все типы питания. Среди них есть всеядные, хищники (рис. 26), растительноядные (рис. 27), детритофаги (сотрирующие и несортирующие) (рис. 28), паразиты, трупоеды (рис. 29) и сестонофаги (рис. 30). Водные растительноядные обычно соскабливают ткани инкрустирующих водорослей с помощью радулы (соскребатели). Детритофаги - собиратели (загребают пищевые частицы). В течение жизни тип питания может меняться (личинка - детритофаг, а взрослый организм - хищник). Хищники либо активно нападают на жертву (могут, например, разбивать или раздавливать раковины двустворок) или протравливают отверстие в наружном скелете жертвы и выедают мягкие ткани. В поиках жертвы гастроподы ползают по поверхности субстрата или зарываются в рыхлый грунт.
Гастроподы чувствительны к изменениям температуры, солености, освещенности и насыщенности вод кислородом, хотя часто один и тот же вид приспосабливается к разным внешним условиям. Температуры существования наземных гастропод от -10 до +50°С, пресноводных от -11 до +56°С, морских от -4 до +36°С. Морские брюхоногие не переносят колебаний солености (существуют в интервале 20-40%о), пресноводны более эвригалинны (0,03-20%о, а некоторые живут и при 150%о). Морские теплолюбивые формы, как правило, более крупные, со сложной скульптурой раковины, пресноводные имеют более тонкую и просто построенную раковину.
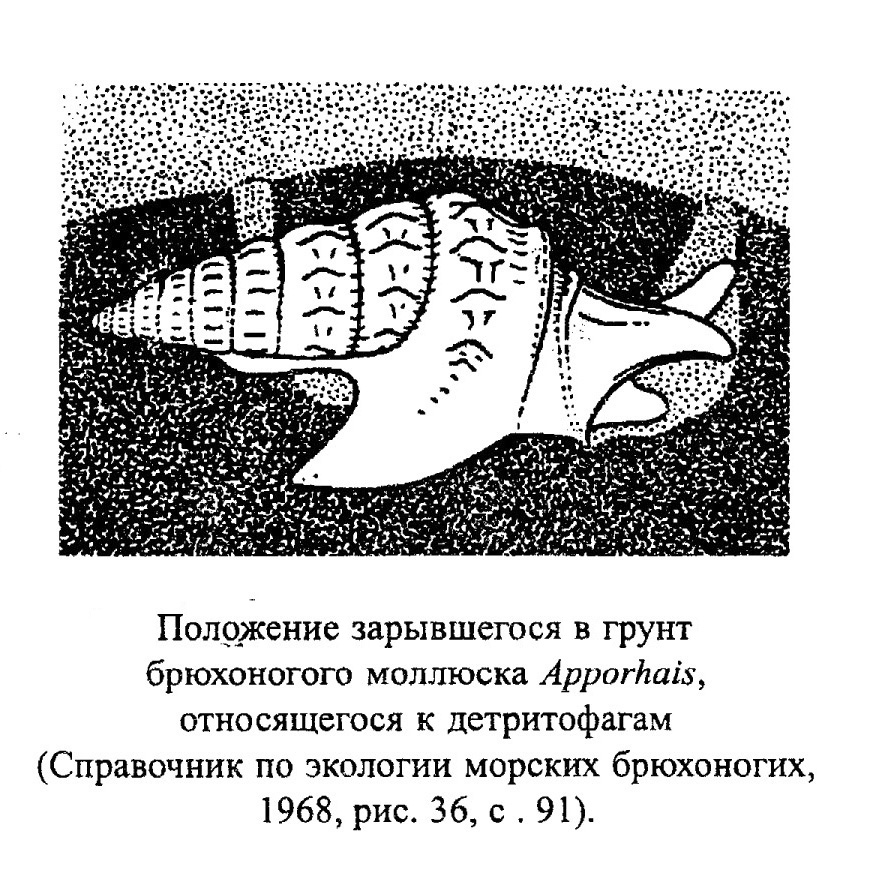
Гастроподы, обитающие в воде, ведут различный образ жизни (ползают или зарываются в донный осадок в поисках пищи, прикрепляются с помощью цемента, плавают). Группы, ведущие сходный образ жизни, обычно имеют близкую морфологию раковин. Для ползающих брюхоногих моллюсков (а самое богатое их сообщество развито на песчаных грунтах) характерна уплощенная нижняя поверхность раковины, они могут также иметь крыловидный край раковины (губу) или шипы для увеличения площади опоры. Растительноядные соскребатели (например, Patella, обр. 385-83, Pleurotomaria, Haliotis, обр. 385-82, Littorina, Trochus, обр. 385-80) могут иметь уплощенную раковину или уплощенную ее нижнюю поверхность. Такую же форму имеет Cyprea, которая соскребает водоросли и объедает живые ткани кораллов. Гастроподы, зарывающиеся в грунт в поисках жертвы, чаще обладают гладкой раковиной (Natica, обр. 385-91) или имеют шипы, расположенные в одной плоскости, которые позволяют им удерживаться на определенной глубине в осадке (Apporhais). У типичных детритофагов Cerithidae (одно из самых распространенных семейств гастропод, живущих на рыхлых грунтах в мелководных зонах тропических морей) - удлиненные толстостенные, часто орнаментированные шипами и выступами раковины. Фильтраторы (сестонофаги), представленные специализированной группой цементноприкрепленных Vermetidae, имеют червеобразные раковины, напоминающие трубки червей-серпулид (рис. 30, обр. 385-93). Также как серпулиды, они могут образовывать органогенные постройки в волноприбойной зоне (рис. 31).
В мезозое и кайнозое при массовом скоплении раковин гастропод часто образовывались ракушняки (обр. 385-94).

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}