Экспозиция "Ископаемые организмы как индикаторы условий осадконакопления"
3. Освещенность
Помимо глубины бассейна, освещенность его зависит от количества илистой взвеси в воде. Глубина проникновения света влияет на расселение донной растительности. Из-за разного количества взвеси в воде нижняя граница распространения донной флоры в различных морях находится на неодинаковой глубине (например, в Балтийском море она не превышает 20 м, а в Средиземном - 160 м).
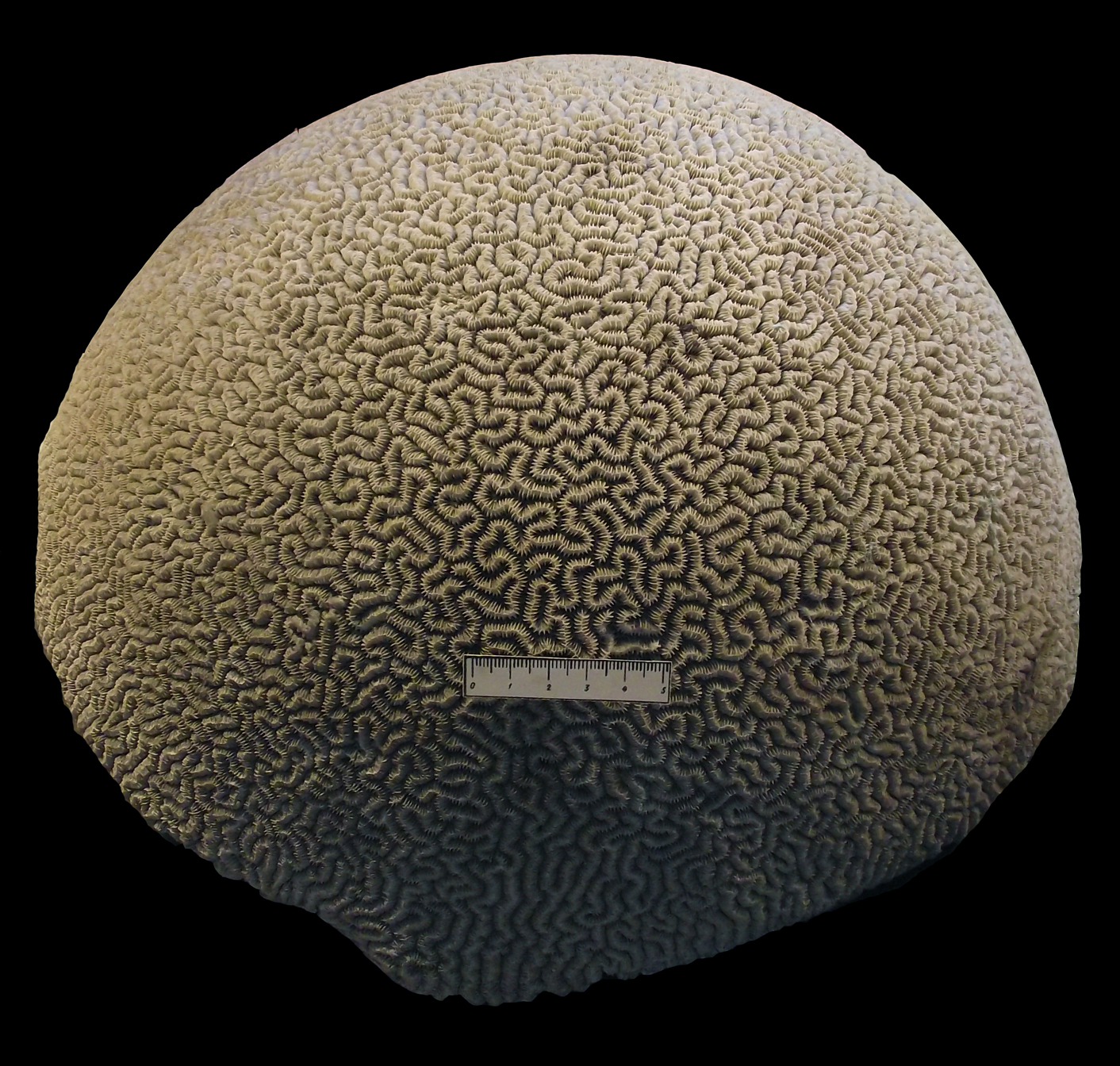
О прозрачности воды в древнем бассейне может свидетельствовать присутствие в породах скелетов рифостроящих кораллов, которые содержали в мягких тканях симбиотические водоросли (обр. 388-6), а также остатков известковых бентосных водорослей и растительноядных животных (гастропод, рыб и др.).
4. Соленость
Общая концентрация солей в океанических водах составляет приблизительно 3,5% или 35%о при постоянном соотношении между главными ионами, к которым относятся ионы хлора, натрия, магния, кальция, калия, сульфат-ион и некоторые другие.
Соленость по большей части не зависит от глубины бассейна или типа осадков. В наиболее открытых морях она почти постоянна (35%о), в прибрежных районах шельфа, в изолированных, полуизолированных бассейнах и лагунах соленость может значительно отклоняться от этой величины. Так, в большей части Балтийского моря соленость составляет 10%о, тогда как в бассейнах, где образуются эвапориты, она достигает 40%о и более.
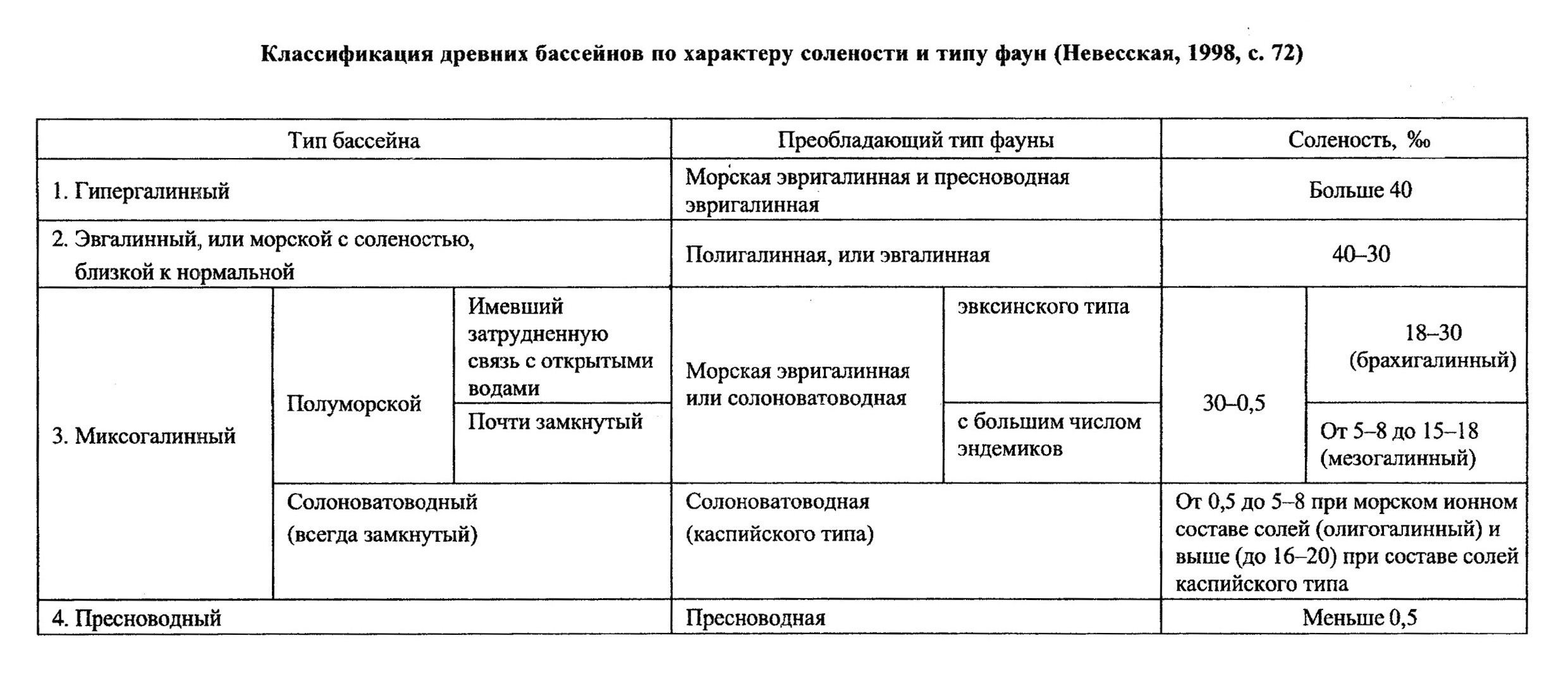
Классификации вод по солености, принятые в настоящее время большинством исследователей, приведены в табл. 1, 2. Есть классификации и более простые, по которым выделяются воды морские, солоноватые и пресные. Границы солености между ними проводятся разными авторами по-разному. Граница морских и солоноватых вод проводится по значению 16%
о (по другим данным - 24,7%
о), граница солоноватых и пресных - по значению 0,5%
о.
Полагают, что в докембрии геохимические особенности океанов и морей значительно отличались от современных. В фанерозое солевой состав Мирового океана был уже близок к современому и менялся мало. Только во второй половине перми соленость океана понизилась до 30-31,5%о, а в эпиконтинентальных бассейнах была еще более низкой.
Для реконструкции солености древних бассейнов применяют физические и геохимические методы (изотопные), данные литологического анализа (по присутствию в отложениях эвапоритов) и анализ палеонтологических остатков.
По отношению к солености организмы могут быть эвригалинными, т. е. живущими в широком диапазоне солености, и стеногалинными, т. е. существующими только в узком ее интервале. К последней группе относятся и солоноватоводные формы, живущие в интервале солености от 5-8%о до 12-14%о. При колебании солености (как повышении, так и понижении) выживают только наиболее приспособленные к этому (эвригалинные) организмы. Морские организмы в большинстве своем плохо переносят эти колебания (то есть являются в основном стеногалинными). Нижняя критическая соленость для морских организмов 5-8%о, верхняя - 42-45%о.
С понижением солености связано уменьшение размеров многих организмов (брюхоногих, двустворок, рыб, иглокожих, некоторых ракообразных). Это происходит в основном с организмами, которым требуется известь для построения скелета. При этом раковины многих моллюсков делаются очень тонкими и утрачивают скульптуру, характерную для особей тех же видов, но обитающих в условиях, где соленость бассейнов близка к нормальной (обр. 388-12).
Резкое опреснение или засоление бассейна может привести к массовой гибели его обитателей (замору). Следы таких событий в геологическом прошлом можно наблюдать в разрезах всего фанерозоя.
Для определения солености древних бассейнов оценивают количество родов, принадлежащих к группам, обитающим преимущественно в бассейнах нормальной солености (фораминиферы, кораллы, цефалоподы, серпулиды, иглокожие, брахиоподы) (обр. 385-21б, 388-6, 388-16, 388-17, 388-19).
С понижением солености общее число родов явственно уменьшается, а видообразование, индивидуальная изменчивость и степень эндемизма организмов повышаются. Чем большему опреснению подвергается тот или иной район моря, тем меньшее число морских форм мы в нем находим (обр. 388-9, 388-13). В наиболее опресненные участки морей проникают только немногие, наиболее эвригалинные морские формы, доходящие иногда до пресной воды. Это можно проиллюстрировать следующими цифрами. В Белом море уменьшение солености на 7-8%
о приводит к обеднению фауны на 50%
, в Черном море уменьшение солености на 18%
о ведет к обеднению состава фауны на 80%
, в Балтийском и Азовском морях при уменьшении солености на 26-27%
о фауна беднеет на 94-95%
.
В лагунах с повышенной соленостью бентос также обеднен. В таких бассейнах часто присутствуют маты из синезеленых водорослей (обр. 386-72).
Большинство обитателей пресноводных бассейнов стеногалинны. Однако и среди пресноводных есть эвригалинные, которые могут встречаться в осолоненных бассейнах совместно с морскими эвригалинными видами (обр. 388-10).
При анализе органических остатков с целью восстановления солености палеобассейнов следует иметь в виду, что он дает возможность устанавливать абсолютную соленость только для очень молодых бассейнов, в которых обитали те же виды, что теперь. Для древних же удается говорить лишь об относительной солености.
5. Газовый режим
Содержание кислорода и углекислоты в океане связано с содержанием их в атмосфере, с характером климата, с распределением континентов, вулканической деятельностью и особенностями океанической циркуляции.
О характере газового режима палеобассейнов можно судить как по литологическому составу отложений, так и по составу заключенных в них органических остатков.
В отдельные промежутки времени в геологическом прошлом дефицит кислорода имел широкое распространение (бескислородные события, аноксия), захватывая почти всю область шельфа.
Литологическим признаком недостатка кислорода в придонном слое воды и на границе воды и осадка обычно служит присутствие черных глин, богатых органическим веществом, которые в процессе диагенеза превращаются в черные сланцы и черные микритовые известняки (как правило, с характерной тонкой слоистостью и мелкими кристалликами пирита, расположенными по плоскостям наслоения).
Из палеонтологических признаков свидетельством нормального снабжения кислородом служит разнообразие бентоса при доминировании сестонофагов, в том числе инфаунных (обр. 385-73, 385-75), и хорошо развитая биотурбация (обр. 390-26). При дефиците кислорода бентос резко обедняется, исчезают инфаунные сестонофаги, почти отсутствует биотурбация, в сообществе преобладают детритофаги (обр. 386-66). Дальнейшее уменьшение содержания кислорода вызывает исчезновение и этой группы организмов. В случае возникновения бескислородных условий с поверхности осадка исчезает весь бентос. В таких условиях могут существовать только представители эпифауны, возвышающиеся над грунтом, то есть находящиеся выше пограничного слоя воды, лишенного кислорода. К ним относятся такие эпибионты, как серпулиды, двустворки, беззамковые брахиоподы, которые жили, прикрепившись к раковинам отмерших головоногих, опустившихся на дно, или же к раковинам погибших бентосных моллюсков. При сильном заражении исчезали и возвышавшиеся над субстратом эпифаунные сестонофаги, так что в анаэробной среде "скелетный" бентос вообще отсутствовал.
6. Гидродинамика
Как и в современных морях, в палеобассейнах гидродинамика оказывала влияние на распределение температур и солености вод, определяла газовый режим, влияла на характер грунтов, прозрачность вод и наличие пищи.
Степень подвижности вод древних морей реконструируется как по литологическому анализу отложений, так и на основании палеонтологических данных.
Так, о наличии сильных волнений или течений в области накопления осадков свидетельствует доминирование грубообломочных и грубозернистых отложений. При этом в зоне активной гидродинамики скелеты бентосных организмов, как правило, быстро распадаются, а фрагменты их хорошо отсортированы (обр. 388-11). В приливно-отливной области часто формировались "ракушечные мостовые" (обр. 388-12). В зоне постоянного действия волн частицы грунта и скелетные обломки хорошо отсортированы и окатаны - здесь формируются биокластические известняки и песчаники с примесью раковинного детрита (обр. 386-22). Организмы в прижизненном положении здесь не сохраняются, целых раковин очень мало, в основном преобладают разобщенные створки.
Высокая степень подвижности воды обусловливает хорошую аэрацию придонных вод и нормальное содержание в них кислорода, в то же время она препятствует отложению осадков и обнажает уплотненное дно. В таких условиях могут существовать только те донные организмы, которые в состоянии противодействовать переносу движением воды. Это животные, которые способны прочно прикрепляться к субстрату, всверливаться в него, а также организмы, обладающие толстостенной раковиной или массивным скелетом другого строения (многие кораллы, мшанки и др.) (обр. 385-43, 385-62, 385-69, 388-1). Высокая подвижность воды приводит к качественной бедности биоценозов, способных существовать в таких условиях. Здесь преобладают сестонофаги, причем в максимально подвижных водах - прикрепленные.
Штормовые воздействия приводили к возникновению специфических осадков - темпеститов, имеющих различный состав на разных глубинах. На глубинах выше базиса штормовых волн формировались раковинные скопления и ил, ниже базиса штормовых волн - тонкозернистые известняки и илы.
В обстановке пониженной гидродинамики преобладают тонкозернистые отложения, а среди бентоса господствуют детритофаги (обр. 385-120).
Гидродинамику палеобассейна с большой достоверностью можно восстанавливать по форме скелетов колониальных организмов (кораллов, строматопорат, хететид, мшанок, известьвыделяющих водорослей и др.). В условиях спокойной гидродинамики в их сообществах преобладают хрупкие ветвистые (обр. 386-43, 386-49, 387-115в) и стелющиеся колонии, а при сильном гидродинамическом воздействии - массивные уплощенные (обр. 385-41) и инкрустирующие формы.
Существование течений в палеобассейнах восстанавливается по присутствию грубозернистых отложений среди тонкозернистых, по наличию косой слоистости, а также по развитию участков с разнообразным и многочисленным прикрепленным бентосом, который жил в условиях высокой гидродинамики. Такие участки располагаются полосами среди отложений, образовавшихся в более спокойной обстановке. Направление течений может быть реконструировано по преобладающей ориентировке удлиненных органических остатков (ростров белемнитов, башенковидных раковин гастропод, скелетов тентакулитов и др.) (обр. 388-18, 388-19, 388-20).
7. Характер грунта
Характер грунта определяется широтой, на которой расположен палеобассейн, глубиной образования осадка, удаленностью зоны осадконакопления от берега, составом материала, приносимого с суши, а также гидродинамикой, скоростью седиментации и некоторыми биологическими факторами (биотурбацией, наличием донной растительности и т. д.). Основными характеристиками грунта как в древних, так и в современных морях являются: степень его стабильности, консолидации, сортировки, аэрации, а также содержание органического вещества, размер зерен и их минералогический состав.
Характер грунта - один из важнейших факторов, определяющих расселение морского бентоса. При этом сами бентосные организмы также оказывают значительное влияние на состояние грунтов. Так, подвижный зарывающийся бентос перемешивает и дестабилизирует грунт, создает ходы, что облегчает циркуляцию растворенных веществ. Сидячие организмы, образующие трубки в грунте, напротив, стабилизируют его. Некоторые организмы пропускают через себя огромное количество осадка, обогащая грунт фекальными пеллетами. Другие выедают из осадка частицы определенного состава и размера, меняя тем самым его структуру. Большое значение имеет наличие отмерших скелетов и их остатков в осадке, так как они могут служить субстратом для прикрепления эпибионтных форм, убежищем для них или объектом для сверления.
Характер грунта древних бассейнов реконструируется с помощью комплекса методов литологического и палеоэкологического анализа. При этом выделяя вертикальные зоны в палеобассейнах, не всегда применяют подразделение на зоны современных морей (литораль, верхняя и нижняя сублитораль, псевдоабиссаль, батиаль, абиссаль), довольно точно удается ихопределить только для морей кайнозоя и мезозоя. Это связано с тем, что данных о составе осадков и характере ориктоценозов бывает недостаточно для того, чтобы точно реконструировать глубины, удаление от берега и другие параметры, по которым устанавливают вертикальную зональность современных бассейнов. Для морей палеозоя чаще выделяют такие зоны (от берега к открытому шельфу), как прибрежное мелководье, удаленное от берега мелководье, или мелководный шельф, и относительно глубоководную зону открытого моря, или глубоководный шельф. В ряде случаев вычленяют еще краевую зону (литораль и лагуны), закрытый шельф, отделенный от открытого мелкого шельфа отмельной зоной, и саму отмельную зону, к которой могут быть приурочены органогенные постройки. Часто шельф как современных, так древних морей подразделяют на внутренний (подверженный волновому воздействию) и внешний (находящийся вне действия волн).
В целом в морях средних и высоких широт преобладали терригенные отложения, а моря низких широт характеризовались широким распространением карбонатов.
Грунты прибрежного мелководья, приблизительно отвечающего литорали, в областях умеренного и холодного климата были терригенными (галечными, песчаными, песчано-глинистыми и глинистыми), в тропической и субтропической областях - преимущественно карбонатными (ракушечными, детритовыми и оолитовыми). Для отложений этой зоны характерно наличие косой и волнистой слоистости, внутриформационных размывов, а также следов волновой ряби и трещин усыхания на поверхностях напластования. Бентос прибрежного мелководья обеднен и представлен наиболее эврибионтными формами, для осадков характерна сильная биотурбация. В результате жизнедеятельности цианобактерий в этой зоне часто образовывались строматолиты.
В обособленных частях морей (лагуны, заливы, бухты) формировались алевролиты, глины, известковые глины, известняки с многочисленными следами илоедов и строматолитами. В гумидной зоне образовывались углистые известняки, в аридной - доломиты, доломитовые известняки, доломитовые мергели.
В тропических и субтропических областях на мелководье формировались рифогенные отложения - разнообразные каркасные известняки с биоморфной слоистостью в органогенных постройках и слоистые органогенно-детритовые известняки в промежутках между ними. Основными каркасообразователями были известьвыделяющие водоросли (обр. 385-15), археоциаты (обр. 385-31), строматопораты (обр. 385-48), кораллы (обр. 387-116), некоторые двустворки (например, рудисты, обр. 385-60), черви-серпулиды (обр. 385-114) и мшанки (обр. 385-111).
На закрытых шельфах отлагались преимущественно илистые осадки, из которых формировались микритовые известняки и известково-доломитовые мергели. Многочисленные цианобактерии образовывали здесь онколиты (обр. 385-10) и сгустки, бентос обеднен.
На мелководном шельфе откладывались преимущественно тонко- и мелкозернистые осадки с параллельной слоистостью - терригенные или карбонатные (в зависимости от климата). С этой зоной связано образование песчаников, алевролитов, детритовых и оолитовых известняков и мергелей. Бентос мелководного шельфа разнообразен, грунты подвергались сильной биотурбации в результате жизнедеятельности зарывающихся организмов (червей, десятиногих раков, двустворок, гастропод и др.) (обр. 386-62).
В области глубоководного шельфа преобладали глинистые, известково-глинистые, карбонатные и кремнистые илы, из которых формировались тонкостоистые глины, аргиллиты, мергели, известковые глины, микритовые известняки, кремнистые глины. В породах хорошо сохраняются остатки планктонных (22), плейстонных (обр. 390-30) и нектонных организмов (24). Бентос этой зоны обеднен и однообразен (особенно при дефиците кислорода), в породах присутствуют характерные следы жизнедеятельности глубоководных животных (обр. 386-61).
С действием апвеллингов в древних бассейнах связывают образование монтмориллонит-глауконит-фосфоритов в ассоциации с организмами, имеющими кремнистый скелет (диатомовые водоросли, радиолярии, губки).
Палентологическими методами можно с большой достоверностью устанавливать степень консолидации осадка. Преобладание таких эпифаунных форм, как кораллы, различные цементноприкрепляющиеся организмы (брахиоподы, гастроподы, двустворки, балянусы и пр.) свидетельствует об уплотненном субстрате (обр. 388-3). Об этом же говорит и наличие сохранившихся постоянных норок глубоко зарывающихся двустворок, ракообразных, иглокожих. Напротив, преобладание инфаунных видов свидетельствует о мягком грунте (обр. 388-21). Бентос на таком грунте, как правило, был обеднен. Известны различные пути адаптации донных организмов к мягким грунтам, в их число входят: уменьшение плотности скелета (образование тонкостенных раковин); погружение в осадок до достижения равновесия между весом организма и весом замещенного осадка (стратегия "айсберга"); увеличение поверхности тела за счет его уплощения (стратегия "лыж") (обр. 385-82); развитие шипов и других выростов (обр. 385-105б); оседание личинок на небольших участках твердого субстрата (например, на обломках раковин) с дальнейшим переходом взрослых форм к свободному лежанию на грунте.
Инфаунные организмы занимали различные уровни в осадке в зависимости от его физических и химических свойств.
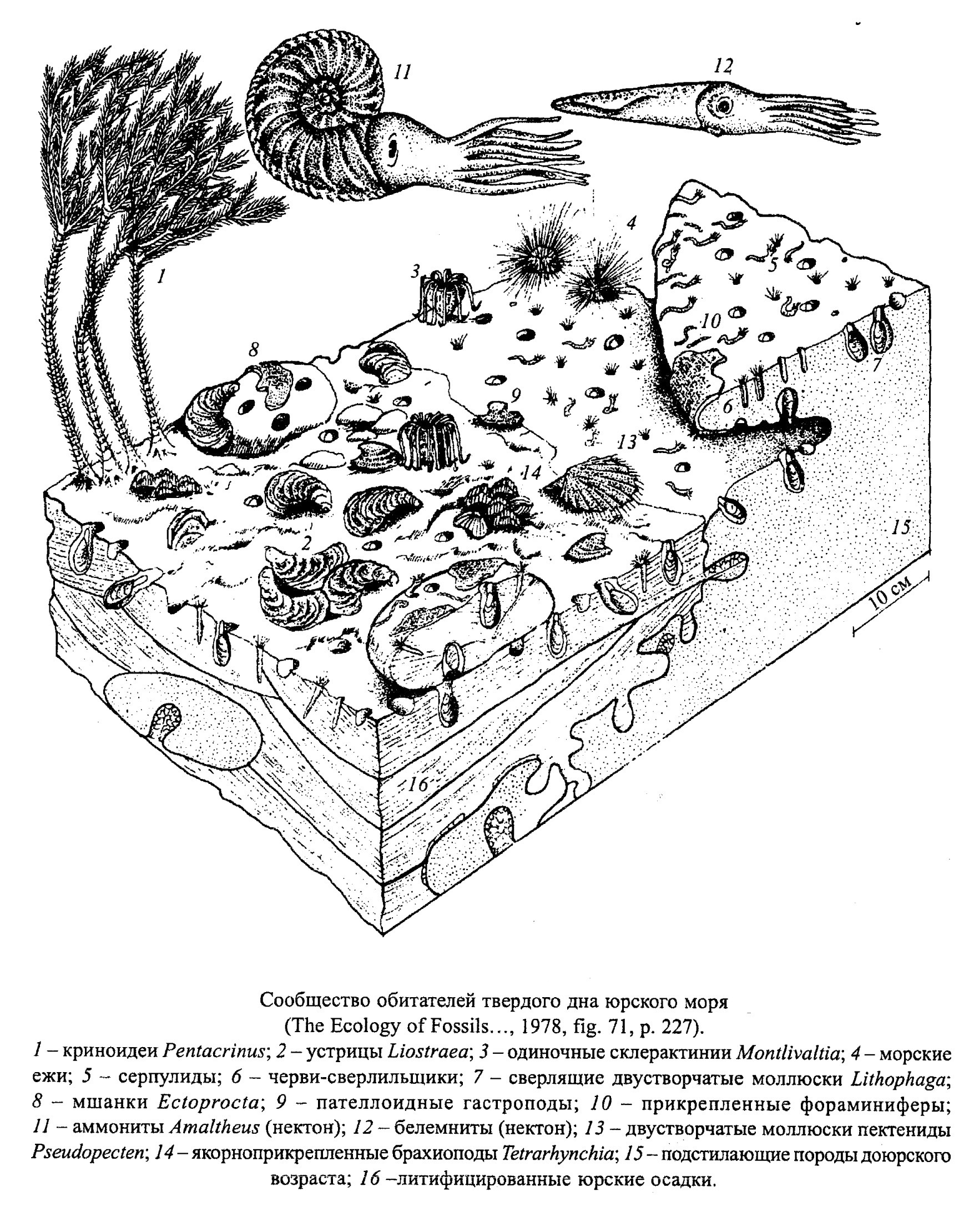
При литификации известковых осадков в подводных условиях на различных глубинах возникает фация твердого дна или хардграунд (hardground) (рис. 10). В древних отложениях ее можно определить по наличию известковых корок, следов карста, по замещению первоначально арагонитовых скелетов кальцитом, а также по следам сверления двустворок, губок, иглокожих, червей и других организмов и присутствию цементноприкрепленных организмов (двустворок, брахиопод, серпулид, мшанок и пр.) (обр. 390-32). Близкий экологический состав имели и скалистые подводные участки. Состав организмов, заселявших твердое дно, зависел от того, на какой глубине оно образовалось.
8. Скорость осадконакопления
Скорость осадконакопления зависит главным образом от интенсивности привноса терригенного вещества в бассейн. В морях с высокой скоростью осадконакопления из донных организмов выживают только подвижные донные эпифаунные и инфаунные животные, а также те из прикрепленных, которые способны занять возвышенное положение по отношению к поверхности дна путем прирастания к выступам дна или скелетным образованиям других организмов (устрицы, брахиоподы и многие другие). Некоторые колониальные организмы (например, рифостроящие кораллы) могут также достигать такого положения за счет высокой скорости скелетообразования, что позволяет им быстро расти в высоту, приподнимаясь над субстратом (обр. 388-5).

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}